
Каковы особенности овуляция яйцеклетки курицы
Опубликовано: 13.04.2024
Процесс яйцеобразования у кур
В связи с интенсивной репродукцией половая система у птицы усиленно функционирует. Она оказывает определенное влияние на форму и функции других органов и систем, испытывая соответственно и их влияние. У птицы действует только левый яичник. Специализация клеток половой системы происходит на первых стадиях развития зародыша. К моменту овуляции пол у птицы уже предопределен. Во время инкубации левый яичник и яйцевод будущих самочек начинают развиваться быстрее, а правый прекращает расти. Например, у эмбрионов кур пол можно различить на 4—5-е сутки инкубации.
При удалении левого яичника в первые дни жизни правый становится семенником и может продуцировать сперму. Однако из-за отсутствия спермиопровода выделение спермы не происходит. При удалении левого яичника в более позднем возрасте правый яичник развивается, могут образовываться фолликулы и даже желтки, но формирование и выделение яйца невозможно. В случаях дегенерации яичника или семенника во взрослом состоянии курица или петух приобретает признаки противоположного пола, включая вторичные половые признаки (особенности оперения, формы гребня, голоса и др.).
Рост яичника и яйцевода до периода полового созревания птицы идет медленно, а в период полового созревания — интенсивно. Если у курочки в возрасте 3 мес яйцевод и яичник имеют массу 0,3—0,5 г, то в возрасте 5 мес — 7—29, а после снесения первого яйца —38—115 г.
Во время яйцекладки яичник у несушек в 10—15 раз больше, чем в период покоя. У курицы в начале первого цикла яйцекладки яичник в 5—7 раз тяжелее, чем при линьке или при прекращении яйцекладки.
В яичнике птицы в фолликулярной зоне наружного (коркового) слоя расположено большое число яйцеклеток. Каждая яйцеклетка находится в отдельном фолликуле, оболочка которого соединена со стромой яичника. Так, в яичнике диких кур и уток насчитывают до 500 видимых невооруженным глазом зачатков яиц — ооцитов; у кур яичного направления — около 3—4 тыс., а при микроскопическом исследовании — до 12 тыс. Чем больше таких зачатков, тем выше потенциальная яйценоскость птицы, но ни у диких, ни у домашних видов птицы она никогда полностью не реализуется. Например, максимальная яйценоскость курицы за всю жизнь, по данным С. И. Сметнева (1978), составила 1519 яиц, а по данным С. И. Боголюбского (1991) — 2036. Задача ученых и практиков состоит в том, чтобы до минимума сократить различия между потенциальной и реальной плодовитостью.
Первичные фолликулы представляют собой по структуре яйцеклетку без желтка. Во вторичных фолликулах желток постепенно накапливается за счет питательных веществ, поступающих через кровеносную систему. Фолликулы увеличиваются не одновременно, а один за другим, достигая размера желтка яйца. У птицы, относящейся к породам, отличающимся высокой яйценоскостью, например у кур яичных пород и уток хаки-кемпбелл, в яичнике больше зрелых фолликулов, чем у одновозрастных с ними кур и уток мясных пород.
Ооцит в организме птицы растет довольно медленно. Усиленно развивается он лишь за 9 сут до овуляции, причем в последние 6 сут диаметр ооцита увеличивается примерно в 5—6 раз. При нарушении условий кормления и содержания птицы значительно снижается скорость образования яиц, а следовательно, и яйценоскость. Возможна даже дегенерация созревающих яйцеклеток. Восстановление нормального процесса формирования яиц и яйцекладки, наоборот, требует некоторого времени. Поэтому необходимо постоянно поддерживать оптимальные условия внешней среды, соответствующие требованиям организма птицы для сохранения высокой продуктивности.
В процессе образования яйца по мере увеличения массы желтка относительное содержание воды в нем уменьшается. Яйцо обогащается жирами, протеинами, минеральными веществами и витаминами.
Окраска желтка в значительной степени обусловлена поступающими с кровью пигментами: каротиноидами, особенно ксантофиллом и каротином. При скармливании птице кормов, богатых каротиноидами (травяная мука, зелень и др.), окраска желтка более интенсивная. Каротин и криптоксантин (представитель группы ксантофиллов) действуют, как провитамин А. По окраске желтка можно судить о содержании в нем каротиноидов, а следовательно, и о витаминной ценности яйца, характеризующей его пищевые и инкубационные качества.
Процессы роста яйца в яичнике находятся под влиянием гормонов и регулируются нервной системой (табл. 3.2). Исследованиями установлено, что ведущую роль в яйцеобразовании играет система: среда—рецепторы—кора головного мозга—гипоталамус— гипофиз—яичник—яйцевод. Гипоталамус при участии гипофиза регулирует температуру тела, содержание воды и крови в тканях, расход углеводов, белков, жиров и минеральных солей, ритм сердца и состояние сосудов.
Гормоны, влияющие на формирование яиц и сезоннуюизменчивостьяйценоскости
Введение в организм гормонов гипофиза способствует быстрому и одновременному увеличению размеров и массы нескольких яйцеклеток. Воздействие света и некоторых других факторов внешней среды стимулирует обмен веществ птицы, ускоряет образование желтков и, следовательно, повышает яйценоскость.
По данным Б. Г. Новикова, ритм работы гипоталамуса, секретирующего рилизинг-гормоны, в конечном счете определяет ритм формирования яиц и сезонную изменчивость яйценоскости. Аденогипофиз (передняя доля гипофиза) выделяет гормоны (фолликулостимулирующий, лютеинизирующий, пролактин, соматотропный, тиреотропный, адренокортикотропный), оказывающие наиболее активное влияние на деятельность половых желез. Нейрогипофиз (задняя доля гипофиза) выделяет три гормона (аргинин-ваз от, окситоцин и антидиуретин), влияющих на процессы яйцеобразования и снесения яиц.
Кроме того, следует особо подчеркнуть, что под воздействием гормонов гипофиза яичник сам становится секреторным органом и выделяет свои гормоны.
В последнюю фазу роста яйцеклетки на поверхности желтка под фолликулярной оболочкой формируется эластичная желточная оболочка, через которую питательные вещества продолжают поступать в яйцеклетку. Когда желток достигает в диаметре 35 — 40 мм, происходит овуляция. Желток освобождается из фолликула вследствие разрыва оболочки последнего вдоль белой линии или вдоль рубчика (истонченная часть фолликулярной оболочки, обращенная в полость тела).
После овуляции кровеносные сосуды фолликула сжимаются и приток крови значительно сокращается. Этим, по-видимому, объясняется отсутствие кровотечения в овулировавшем фолликуле. В то же время происходят морфофизиологические изменения в кровеносной системе, ведущие к усилению кровоснабжения другого очередного фолликула, подготавливающегося к овуляции.
Под влиянием нейрогуморальных факторов овулировавшая яйцеклетка с большим запасом питательных веществ попадает в воронку яйцевода, прилегающую к яичнику. Здесь при наличии достаточного числа жизнеспособных спермиев происходит ее оплодотворение.
Яйцевод птицы представляет собой относительно длинную извилистую, очень эластичную трубку, передний конец которой открывается в полость тела вблизи яичника, а другой — в клоаку. Диаметр яйцевода при прохождении яйца увеличивается. Брыжейка, на которой подвешен яйцевод, допускает значительные его движения. В зависимости от физиологического состояния и продуктивности птицы размер и масса яйцевода сильно изменяются. Так, у несущейся курицы длина яйцевода около 15 см, во время интенсивной яйцекладки — 75 см и более, а ширина увеличивается от 0,5 до 10 см.
В яйцеводе различают воронку, белковую часть, перешеек, матку и влагалище. У курицы, прекратившей яйцекладку, воронка, матка и влагалище уменьшаются в 2,5—3,5 раза, а белковая часть — в 6 раз.
Стенка яйцевода состоит из наружной серозной оболочки, продольных мышц; соединительной ткани с большим количеством кровеносных сосудов; кольцевых мышц; слизистой оболочки (с интенсивно развитыми кровеносными сосудами и железами), образующей мелкие и большие складки. Слизистая оболочка покрыта реснитчатым эпителием. Секреторную функцию яйцевода выполняют клетки эпителия и трубчатых желез слизистой оболочки.
За счет перистальтических движений стенок яйцевода и их складчатости яйцо совершает вращательное движение вдоль продольной оси. Вокруг желтка яйца прежде всего наслаивается наружный плотный белок, состоящий из тончайшей сети волокон муцина. Плотный белок у острого и тупого концов яйца образует спиралеобразные градинки (халазы), которые удерживают желток в центре яйца. При дальнейшем движении яйца по яйцеводу появляется слой среднего плотного белка. Между ними постепенно накапливается внутренний жидкий белок из секрета трубчатых желез слизистой оболочки яйцевода. Вращение яйца приводит к выделению жидкого белка внутрь плотного.
В белковом отделе яйцевода образуется 40—50 % белка, а остальная часть секретируется в перешейке и матке.
В перешейке яйцевода формируются белковая и подскорлуп-ная оболочки, состоящие в основном из белка, выделяемого железами этой части яйцевода; кроме того, в яйцо продолжают поступать растворимые в воде неорганические вещества.
В матке яйцевода образуется скорлупа и, следовательно, выделяется большое количество минеральных веществ, главным образом кальция (около 5 г за 20 ч) и фосфора. Перед яйцекладкой и во время ее содержание кальция в крови несушек возрастает в 2— 3 раза; увеличивается и количество фосфора. При недостатке кальция в корме организм птицы мобилизует его из костяка. Если этого оказывается недостаточно, то птица несет яйца без скорлупы («литые яйца»). В результате нарушения обмена веществ яйцекладка может прекратиться.
При образовании скорлупы сначала на поверхности яйца появляются лишь отдельные отложения кальция, которые постепенно увеличиваются. Среди них находятся небольшие количества органических веществ в основном белкового характера. Это способствует образованию сосочкового слоя скорлупы. Основание каждого сосочка связано с подскорлупной оболочкой. Сосочки постепенно увеличиваются, боковые стенки их соприкасаются друг с другом, но между ними остаются небольшие поры, через которые в яйцо проникает воздух. Железами передней части матки выделяется протеин, который в виде волокон располагается на сосочковом слое матки.
Пространство между сетью протеиновых волокон заполняется кристаллами выделяющихся солей кальция, что в конечном итоге приводит к образованию плотного и крепкого губчатого слоя скорлупы. Причем губчатая структура слоя становится заметной лишь при удалении солей кальция. Готовое к снесению яйцо через влагалище выталкивается наружу.
Проходя по яйцеводу, формирующееся яйцо растягивает стенки яйцевода, создавая этим биоэлектрический потенциал, стимулирующий синтез простагландинов, которые вместе с аргинин-ва-зотоцином и прогестероном вызывают сокращение гладких мышц яйцевода и перемещение образующегося яйца на новый участок полового пути.
Скорость формирования яйца у птицы разных видов, а также у низко- и высокопродуктивных особей различна. У высокопродуктивных, ежедневно несущихся кур средняя продолжительность формирования яйца составляет 24—25 ч (табл. 3.3). У хороших несушек примерно через 30—40 мин после снесения яйца наступает новая овуляция, которая проходит в основном в период от 6 до 15 ч дня.
Продолжительность формирования яйца в яйцеводе курицы (по С. И. Боголюбскому, 1991)
Отмечена возможность формирования нормальных яиц у кур за 18—21 ч (М. Е. Лобашов, A. JI. Штеле). Путем селекции удалось уменьшить срок формирования яиц на 3 ч 53 мин. Наследуемость данного признака высокая (0,66—0,75), но быстро уменьшающаяся с возрастом птицы.
Сокращение времени образования яйца в яйцеводе связано в основном с более ранним началом формирования скорлупы и большей скоростью ее минерализации, которая определяется интенсивностью обмена кальция в организме и содержанием его в крови. Уровень кальция в сыворотке крови у высокопродуктивных несушек составляет 24—26 мг%.
Установлено, что чем меньше времени затрачивает несушка на формирование яйца, тем длиннее у нее циклы (серии) яйценоскости. Циклом яйценоскости называют число яиц, снесенных несушкой без интервала, то есть подряд. Длина циклов — наследуемая особенность птицы. Циклы могут составлять от одного до нескольких десятков яиц. Между циклами образуются интервалы, выражаемые числом непродуктивных дней. Чем меньше цикл, тем длиннее интервал, и наоборот. Длинные циклы с короткими интервалами характеризуют хороших несушек, короткие циклы с длинными интервалами — плохих. Во время длинных циклов снесение яиц происходит почти в одни и те же часы, за исключением нескольких дней в начале и в конце цикла. Отмечены случаи снесения курицей двух яиц в сутки.
Классическим объектом эмбриологических исследований является куриный эмбрион. Яйцеклетка у птиц богата желтком — полилецитальная. Диаметр яйцеклетки курицы достигает 35 мм, а у страуса — до 100 мм. В отложенном птичьем яйце находится не яйцеклетка, а многоклеточный зародыш на стадии бластулы или ранней гаструлы. К яйцеклетке относится то, что мы называем желтком. Яйцеклетка, продвигаясь по яйцеводу, оплодотворяется. Диаметр овулировавшего яйца составляет 50 мкм и постепенно увеличивается до 6 мм, а затем и до 35 мм.
Желточный материал синтезируется в печени, переносится в фолликулярные клетки, а затем в цитоплазму яйцеклетки. У рептилий и птиц наблюдается полиспермия и, хотя с женским пронуклеусом сливается один мужской, избыточные спермин — мероциты (блуждающие ядра) способствуют переработке желтка.
Дробление зиготы птиц неполное, неравномерное, частичное, дискоидальное, так как дробится всего 1/500 часть зиготы на анимальном полюсе. Первые три борозды дробления — радиальные, затем появляются широтные и тангенциальные полосы. При дроблении возникают бластомеры различной величины, и в конечном итоге формируется многослойная дискобластула, распластанная на нераздробившемся желтке.
Как отмечалось выше, к моменту откладки яйца зародыш находится в первой фазе гаструляции. Это проявляется обособлением внезародышевой энтодермы путем деламинации дискобластулы и возникновением двуслойного зародыша. Так возникают две части: эпибласт — наружная и гипобласт — внутренняя части дискобластулы (рис. 28). Периферическая часть гипобласта в дальнейшем образует внезародышевую энтодерму, которая обрастает желток, формируя желточный мешок — провизорный (временный) орган, выполняющий трофическую и кроветворную функции. В стенке желточного мешка обнаруживаются первичные половые клетки (внегонадное происхождение).
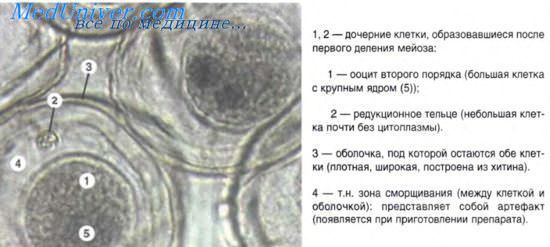
Наиболее важные морфогенетические процессы развертываются в эпибласте — источнике развития всех трех зародышевых листков. Здесь возникают клеточные потоки вследствие асинхронного дробления клеток и их направленного перемещения. Клетки перемещаются в течение 3-4 часов инкубации. Схема движения клеточных потоков выглядит следующим образом: клетки латеральных частей эпибласта перемещаются в хвостовую часть зародыша и, встречаясь здесь, движутся далее по средней линии эпибласта к головному концу. К 12 часам клеточный поток принимает вид первичной полоски.
Второй поток клеток (более медленный) распространяется со стороны головного конца зародыша и движется по средней части эпибласта навстречу первому потоку. Место встречи двух потоков обозначается как гензеновский, или первичный, узелок — важнейший гистологический маркер. На вершине гензеновского узелка появляется углубление — первичная ямка, а по средней линии первичной полоски — первичная бороздка, как продолжение первичной ямки. Обе эти структуры рассматриваются как гомолог бластопора. К 16 часам инкубации все описанные структуры хорошо выявляюся на фиксированных препаратах.
Дальнейшие перемещения клеток — иммиграция, характеризуют вторую фазу гаструляции. В области переднего края гензеновского узелка клетки, подворачиваясь, проходят через ямку и распространяются в направлении головного конца зародыша, формируя головной отросток — зачаток хорды (нотохорд). В свою очередь, зачаток хорды индуцирует раннюю детерминацию клеток нервной пластинки в составе эпибласта. Одновременно с материалом хорды из противоположного края гензеновского узелка и передней части первичной полоски в иммиграцию вовлекаются новые клетки, однако последние включаются в состав гипобласта головной части зародыша.
При этом происходит деформация гензеновского узелка и укорочение первичной полоски. Все эти процессы протекают в течение 8-10 часов инкубации. Клетки оставшейся части первичной полоски — представляют собой материал сомато- и спланхномезодермы. Через 15 часов инкубации эти клетки приходят в движение и мигрируют внутрь вдоль первичной полоски, распространяясь над гипобластом по бокам от удлиняющейся хорды.
Таким образом, в результате иммиграции из эпибласта выселяются материал хорды, прехордальной пластинки (а, по некоторым данным, материал зародышевой энтодермы), мезодермы, что сопровождается деформацией, укорочением и исчезновением гензеновского узелка и пер вичной полоски. Оставшийся материал эпибласта представляет собой эктодерму, которая в своем составе содержит клетки будущей нервной трубки. Хорда индуцирует развитие нервного желобка, сворачивающегося затем в трубку и погружающегося под эктодерму.
Клеточный материал мезодермы дифференцируется на сомиты, латерально расположенные нефротомы и наиболее латерально лежащие спланхнотомы. Сомиты и нефротомы — это сегментированные части мезодермы, спланхнотом — несегментированная. Последний в дальнейшем расслаивается на висцеральный и париетальный листки со щелевидной целомической полостью между ними. Под хордой располагается энтодерма, которая сворачивается в кишечную трубку, сообщающуюся с желтком. Так возникают эмбриональные зачатки органов и тканей. Наиболее сложна топографическая дифференцировка мезодермы. В составе сомита различают следующие части: склеротом — вентромедиальную часть, дерматом — дорсолатеральную часть и миотом — дорсомедиальную часть.
Клетки склеротома интенсивно размножаются, выселяются из состава сомита, окружают хорду и дифференцируются в хрящевые клетки (возникают сложные формообразовательные процессы закладки позвонков, ребер, лопаток и других костей скелета). Дерматом является источником развития соединительных тканей кожи, а миотом — источником развития скелетной мышечной ткани.
Параллельно образованию тела зародыша происходит образование внезародышевых оболочек, или провизорных (временных) органов. У зародышей высших позвоночных имеются 4 вида оболочек — амнион (водная), хорион (серозная оболочка), аллантоис и желточный мешок.
Изучение процесса формирования и оплодотворения куриного яйца является необходимой предпосылкой для выработки правильного представления об эмбриогенезе цыпленка.
Это тем более понятно, что большая часть развития цыпленка до его выклева из яйцевой скорлупы происходит вне непосредственной связи с материнским организмом, осуществляясь за счет тех веществ, которые окажутся включенными в состав отложенного курицей яйца.
Можно назвать большое количество работ, авторы которых изучали различные стороны процесса оогенеза, овуляции и оплодотворения куриного яйца. Наряду с ними имеются исследования и по изучению дробления яйца курицы и по ранним стадиям развития зародыша, протекающих до начала инкубации. Однако подавляющее большинство литературных данных по этим важным разделам раннего онтогенеза кур представлено в виде узкоспециальных исследований, причем выводы авторов по отдельным вопросам иногда носят противоречивый характер. Сколько-нибудь удовлетворительных сводок на русском языке по указанным проблемам нет.
Исходя из этого, нам представляется целесообразным кратко резюмировать ранее полученные данные по каждому из затронутых разделов и отметить те спорные положения среди них, которые еще ждут своего разрешения.
Оогенез. В оогенезе птиц, как и в оогенезе других животных, различают три периода: период размножения яйцевых клеток, период их роста, подразделяемый в свою очередь на период малого и большого роста, и период их созревания. Период размножения оогоний, происходит еще в яичнике зародыша (Де Голандер — D’Hollander, 1905, Венцке — Venzke, 1954). Яйцевые клетки в этом периоде относительно малы. Их диаметр не превышает 0,016 мм (Свифт — Swift, 1914). Период размножения половых клеток, темп деления которых велик, заканчивается ко времени вылупления цыпленка из яйцевых оболочек.
В период малого роста половые клетки или ооциты начинают увеличиваться в размере за счет образования и накопления в них первичного желтка. К этому времени они окружаются однослойным фолликулярным эпителием яичника. По данным Брэмбел (Brambell, 1926), клетки фолликулярного эпителия выделяют в цитоплазму ооцита тельца Гольджи, которые в дальнейшем становятся центрами образования желтка.
В период большого роста в плазме яйца, в результате постепенно усложняющейся секреторной деятельности фолликулярного эпителия яичника (Мазунин, 1955), начинает накапливаться вторичный желток. Процесс развития ооцитов сравнительно длителен. В нем различают три следующие друг за другом фазы желткообразования, определяемые как свойством входящего в их состав желтка, так и количеством последнего в ооците (Романов, 1931; Мырза В. и Мырза Е. — Marza Е. a. Marza Е. 1935; Далмане, 1955).
В начальной фазе желткообразования, совпадающей с одновременным ростом ооцита за счет увеличения в нем цитоплазмы, в последней происходит накопление резервных веществ, состоящих из капель нейтрального жира и липоидов. Накопление желтка в ооците цыпленка начинается на 10 день его развития до вылупления из яйцевых оболочек (Гольдсмит — Goldsmith, 1928). В результате этого ооциты увеличиваются в размере. Их диаметры в среднем возрастают от 20 до 700 мк. Клеточное ядро находится в это время в центре ооцита.
Вторая фаза желткообразования характеризуется появлением в резервном веществе ооцита желточных вакуолей (Конопацка — Konopacka, 1933; Далмане, 1955). Она заканчивается формированием первичного желтка более светлого цвета. Отложение первичного желтка начинается в центре ооцита, там, где в дальнейшем образуется латебра яйца. Клеточное ядро вскоре после этого перемещается к периферии ооцита. Оттягивая за собой часть светлого желтка, оно образует при этом шейку латебры. Обычно диаметры ооцитов в это время равны 1,0 мм, тогда как к концу второй фазы желткообразования они в среднем достигают 3,0 мм.
Второй фазой желткообразования заканчивается период малого роста ооцита.
Третья фаза желткообразования, характеризующаяся появлением в ооците вторичного желтка, связана с перестройкой секретирующей деятельности фолликулярного эпителия яичника и, в частности, с проявлением у него способности пропускать в ооцит жиры, растворяющие красители. В связи с этим окраска желтка яйца меняется от светло-желтых тонов до интенсивно оранжевого цвета в зависимости от характера получаемой несушкой пищи.
Шары желтого желтка значительно больше шаров белого желтка. Это в свое время было отмечено еще Бальфуром (Balfour, 1881), который считал, что диаметр первых бывает в пределах от 25 до 100, а вторых от 4 до 75 мк.
Подробно морфологические особенности элементов желтка куриного яйца породы белый леггорн изучены Кнорре (1955). Автор использовал для этой цели морфологические и гистохимические методы исследования с применением фазово-контрастной, ультрафиолетовой и люминесцентной микроскопии. По его данным, желтые желточные шары могут быть мелкозернистыми и грубозернистыми. Их диаметры варьируют от 20 до 60 мк. Так называемые белые желточные шары более мелкие, чем желтые и структура их гомогенна. Их диаметры не превышают 7—20 мк. Автор описывает также ряд промежуточных форм желточных шаров. И белые, и желтые шары, по его данным, содержат нейтральный жир. В желтых шарах жир пропитывает всю его основу, а в белых содержится только по их периферии.
Данные Кнорре совпадают с выводами Датковны (Datkowna, 1949), которая нашла, что к началу инкубации куриного яйца желток представляет собой эмульсию жировых капель, диспергированных в жидкости коллоидного фосфоропротеина.
Третья фаза желткообразования, совпадающая с периодом большого роста ооцита, особенно интенсивно протекает во время яйцекладки. Желток откладывается или непосредственно под фолликулярным эпителием, или под образовавшейся в последней фазе роста ооцита на его поверхности бесклеточной оболочкой или zona radiata.
На более ранние слои желтка наслаиваются последующие его отложения, что может придать сформированному ооциту слоистый вид. Скорость образования желтка нарастает с увеличением поверхности ооцита и является производной роста фолликулярного эпителия. За несколько дней до овуляции темп отложения желтка снижается. По данным Борковской (1954), на образование вторичного желтка затрачивается от 6 до 9 суток и лишь изредка на это требуется до 20 дней.
Zona radiata, окружающая ооцит до его овуляции, представляет собой продукт жизнедеятельности внутреннего слоя клеток фолликулярного эпителия. Таким образом, она по своему происхождению относится к вторичным яйцевым оболочкам. На более поздних стадиях zona radiata сливается с первичной оболочкой яйца, выделяемой цитоплазмой яйцеклетки. После этого оба образования не могут быть разделены.
Zona radiata имеет отверстия в виде многочисленных тонких канальцев, через которые проникают тончайшие отростки протоплазмы клеток фолликулярного эпителия, достигающие своими концами до цитоплазмы ооцита. Этим путем происходит поступление из крови матери в ооцит веществ, используемых на формирование желтка.
К сожалению, среди исследователей нет достаточной ясности в вопросе о том, какое образование в курином яйце следует называть желточной оболочкой. Обычно желточной оболочкой называют zona radiata, плп то более сложное по своему происхождению образование, которое появилось после дополнительного присоединения к ее первоначальной структуре сети муциновых волокон, выделенных из первой порции белковой оболочки и отложившейся на ее поверхности после овуляции яйца и попадания его в воронку яйцевода (Нидхэм и Смит — Needham a. Smith, 1931; Ганзен — Hansen, 1934; Конрад и Филлипс — Conrad a. Phillips, 1938; Нидхэм, 1942 и др.).
Таким образом, в состав того образования, которое обычно принято называть желточной оболочкой яйца, оказываются включенными: первичная или собственно желточная оболочка — продукт цитоплазмы яйцеклетки; вторичная оболочка, или производная клеток фолликулярного эпителия, zona radiata, и незначительная часть третичной оболочки, или халазовая оболочка яйца. Последняя может быть так названа, так как при дальнейшем формировании третичных оболочек из отходящих от нее волокон муцина с двух противоположных концов желточного шара образуются закрученные тяжи, или халазы, одна из которых направлена к тупому, а другая к острому концам яйцевой скорлупы.
В дальнейшем термином «желточная оболочка» нами будет обозначаться общепринятое для этого понятия образование.
О том, что желточная оболочка не представляет собой гомогенной массы, свидетельствуют данные ряда исследователей, изучавших ее структуру. Так, при помощи люминисцентного микроскопа Шалумовичу (1955) удалось различить в ней пять слоев. Однако этот же автор на основе гистологического исследования и ультрафиолетовой микроскопии выявил ъ ней только три слоя. Три слоя в желточной оболочке куриного яйца обнаружили Моран и Холл (Moran a. Hall, 1936) и Мак Нелли (McNally, 1943), которые показали, что ее внутренний слой толщиной от 3 до 6 мк состоит из муцина, средний толщиной от 6 до 10 мк — из кератина и внешний толщиной от 3 до 6 мк, так же как и первый, — из муцина. В поверхностном слое желточной оболочки ими была выявлена отчетливо выраженная волокнистость. Окончательно сформированная желточная оболочка не имеет пор или отверстий, она состоит из переплетающихся волокон кератина и муцина. Вода легко проходит из наружной среды через желточную оболочку в желток.
Процесс созревания куриного яйца подробно описан Олсен (Olsen, 1942, 1951); Олсен и Франс (Olsen a. Fraps, 1944, 1950). По их данным, начало созревания яйца происходит до его овуляции; оно связано с процессом перемещения ядра в ооците из центрального местоположения в нем к периферии. По мере миграции ядра происходит изменение его формы, которая постепенно из шаровидной становится чечевицеобразной. В результате этого процесса на анимальном полюсе яйца из ядра и небольшого количества протоплазмы образуется диск, диаметр которого равен 3,5 мм.
По предположению Борковской (1954), нахождение ядра и протоплазмы на поверхности желточного шара указывает на их связь с фолликулярным эпителием, так как последний, по ее данным, теряет в этом месте способность выделять желток.
Примерно за сутки до момента овуляции в ядре куриного яйца происходит распад ядерной оболочки, т. е. наступает подготовка ядра к делению, в результате которого образуется первое направительное тельце. Первое направительное тельце выделяется из яйцевой клетки примерно за час до овуляции, оставаясь лежать после этого под zona radiata. Веретено второго деления созревания появляется в яйце незадолго до овуляции, однако выделение второго направительного тельца происходит уже после разрыва фолликула, попадания яйца в воронку яйцевода и проникновения в яйцо спермия.
Овуляция и оплодотворение. После достижения яйцевой клеткой максимального размера, вследствие накопления в ее цитоплазме желтка, происходит овуляция. Фолликулярный эпителий в. том месте фолликула, которое носит название стигмы, или рубца, под влиянием мышечного сокращения стенки фолликула, наступающего незадолго до овуляции, постепенно утоньчается, а затем разрывается. Яйцо выпадает из фолликула в просвет воронки яйцевода, охватывающего в это время яичник (Кёртис — Curtis, 1910). Фолликулярный эпителий, плотно сращенный по краю рубца со стенкой яичника и соединенный с ним с внутренней стороны специальной сосудистой ножкой, после выпадения яйца из фолликула остается на месте.
Что вызывает сокращение мышц стенки фолликула, приводящего к разрыву рубца, остается спорным. Предположение Зоненбродт (Sonnenbrodt, 1908) и Паттерсона (Patterson, 1910) о том, что в этом процессе принимают участие мускульные сокращения, производимые воронкой яйцевода, охватывающей фолликул перед его разрывом, не подтвердилось экспериментальной работой Пирль и Кёртиса (Pearl a. Curtis, 1914), удалявших яйцев од и наблюдавших при этом нормальный процесс овуляции. Бартельмец (Bartelmez, 1912) на основании изучения оогенеза у голубя сделал вывод о том, что разрыв рубца происходит в результате механического давления со стороны увеличившегося в объеме желтка на клетки фолликулярного эпителия. Это давление стимулирует фолликулярный эпителий к выделению новых порций желтка, пока увеличение яйца не достигнет такого масштаба, что наступит разрыв рубца в фолликуле.
Очень вероятно, что этот механизм, коррелируемый гормональной «системой курицы, в какой-то мере способствует своевременному процессу овуляции.
Особый интерес представляет вопрос о том, с каким интервалом происходит овуляция у кур в период интенсивной яйцекладки, так как это помогает уточнять истинный возраст зародыша. По данным Филлипс и Уоррен (Phillips a. Warren, 1937), между интервалом в откладке яиц и овуляцией имеется положительная корреляция. При исследовании 84 кур породы белый леггорн, они обнаружили, что в среднем интервал между откладкой следующих друг за другом яиц равен 26,3 часа, с вариациями в пределах от 24,3 до 28,7 час. Интервал между откладкой яйца и новой овуляцией в среднем оказался равным 32,2 мин. с вариациями от 7 до 74 мин. Результаты, близкие к этим данным, были несколько ранее получены Мак Нелли и Байерли (Me Nally a. Byerly, 1935) и Уорреш и Скотт (Warren a. Scott, 1935), что указывает на достоверность наблюдаемого явления.
Чтобы иметь возможность установить возраст зародыша до начала инкубации, определяемый временем нахождения оплодотворенного яйца в половых путях курицы, необходимо предварительно достаточно точно знать, где, т. е. в каком участке яйцевода, происходит процесс оплодотворения.
Проследить проникновение спермиев в яйцеклетку и изучить последующее слияние женского и мужского пронуклеусов в курином яйце оказалось технически трудной задачей, вследствие накопления в плазме яйцеклетки птиц большого количества желтка. Вероятно, этим объясняется получение противоречивых данных по этому вопросу в работах разных исследователей. Иванов (1924) и Войтишкова (Vojtiskova, 1955) считают, что оплодотворение яйца курицы наступает еще до овуляции, т. е. непосредственно в яичнике. Другие авторы: Петтерсон (Patterson, 1910), Уолтон и Уисхем (Walton a. Whetham, 1933) полагают, что оплодотворение куриного яйца наступает в верхних отделах яйцевода, непосредственно вслед за его попаданием туда. Вместе с тем Колъбруге (1913) наблюдал, что сперматозоиды не поднимаются по яйцеводу выше перешейка и на этом основании приходит к выводу, что оплодотворение куриного яйца не может происходить сразу же вслед за овуляцией.
Пересадкой зрелых яйцеклеток от искусственно осемененной курицы в воронку яйцевода другой стерильной курицы Олсен и Нир (Olsen а. Neher, 1948) доказали, что оплодотворение яйца происходит сразу же после его овуляции.
Изучая оплодотворение у кур Бехтина (1955, 1958) установила, что этот процесс происходит в воронке яйцевода и наступает по прошествии 10—30 мин. после овуляции. По ее данным, яйцо к этому времени находится на стадии второго деления созревания.
Подведя итог всем упомянутым работам по выяснению сроков овуляции и оплодотворения куриного яйца, легко обнаружить могущую возникать изменчивость этого процесса. В частности, она может быть продемонстрирована следующим, чисто теоретическим, подсчетом. Предположим, что в некотором стаде куры откладывают яйца с интервалом, изменяющимся от 24 час. 18 мин. до 28 час. 42 мин. Промежутки между откладкой яйца и овуляцией у этих кур будут варьировать в пределах от 7 мин. до одного часа 14 мин. Принимая во внимание, что интенсивность овуляции непосредственно скоррелирована с интенсивностью кладки, будем считать, что у курицы, кладущей яйцо с интервалом в 24 час. 18 мин., овуляция наступит через 7 мин. после откладки предыдущего яйца, а у курицы, кладущей яйца с интервалом в 28 час. 42 мин., овуляция наступит через один час 14 мин. Зная, что оплодотворение яйца происходит в промежутке кежду 10 и 30 мин. после овуляции, можно принять, что в среднем она наступит в обоих случаях через 20 мин. Тогда оплодотворение нового яйца у первой курицы наступит после откладки ею предыдущего яйца через 27 мин., а у второй курицы через час 34 мин. У первой курицы развитие зародыша до откладки яйца будет длиться 23 час. 51 мин. (24 час. 18 мин. минус 27 мин.), а у второй курицы на это пойдет 27 час. 8 мин. (28 час. 42 мин. минус час 34 мин.). Таким образом, зародыш от второй курицы будет ко времени откладки яйца старше зародыша первой курицы на 3 час. 17 мин. (27 час. 8 мин. минус 23 час. 51 мин.). Эти вычисления показывают, что даже от такого в общем однотипного куриного стада предел вариации возрастов зародышей в только что отложенных яйцах составляет более трех часов. Если принять во внимание, что для промышленной инкубации яйца берутся от кур, откладка яиц у которых происходит с значительно отличающихся друг от друга интервалами во времени, то станет понятным, что различия в возрасте зародышей ко времени откладки яиц могут быть более значительными.
В промышленную инкубацию могут попасть яйца с разными сроками хранения после откладки. Как показали исследования Шишкиной (1949), длительное хранение яиц замедляет последующий рост и дифференцировку зародышей. Исходя из указанного выше, всегда следует иметь в виду, что при изучении строения зародышей, взятых из яиц, одновременно заложенных в инкубатор, исследователю заведомо приходится иметь дело с разновозрастными зародышами.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
ИЗУЧЕНИЕ ОРГАНОВ ВОСПРОИЗВОДСТВА С/Х ПТИЦЫ
Цель занятия. Изучить строение и функции органов размножения самок и самцов птицы. Ознакомиться с процессами формирования яйца.
Содержание и методика проведения занятия. У птицы независимо от ее вида половые органы устроены одинаково.
У самцов птицы имеются парные семенники и семяпроводы, расположенные в брюшной полости тела.
Семенники (рис. 44) птицы парные, имеют бобовидную или овальную форму, расположены вдоль позвоночного столба в брюшной полости у переднего края почек.
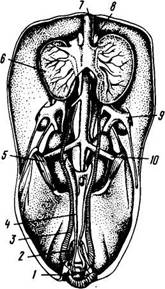
Рис. 44. Половые органы петуха:
1 – клоака; 2 – конец прямой кишки; 3 – семяпровод; 4 – мочеточник;
5 – почка; 6 – семенник; 7 – аорта; 8 – надпочечник; 9 – подвздошная артерия; 10 – крестцовая артерия
У половозрелых петухов длина семенников достигает 4,7 см, ширина - 2,7 см, а масса – 17-19 г. Во время линьки, когда понижается половая активность птицы, масса органа снижается до 3-5 г. Каждый семенник имеет слабо развитый придаток, который переходит в семяпровод.
Первичные половые клетки называют сперматогониями, из них образуются сперматоциты первого и второго порядка, затем сперматиды и спермии. Зрелые спермии через просвет извитых канальцев поступают в семяпровод. В придаток семенника впадает большое количество выносящих канальцев, по которым передвигаются спермин. Выносящие канальца сливаются в проток, переходящий в семяпровод-тонкую извитую трубку, стенки которой состоят из трех оболочек: слизистой, мышечной и серозной. На конце семяпровода имеется расширение для накопления спермы. В клоаку семяпровод открывается маленькими сосочками около отверстия мочеточника.
Копулятивный орган у петухов и индюков не развит. При спаривании самец прижимается клоакой к клоаке самки, происходит выпячивание влагалища и сперма попадает в половые пути самки, минуя клоаку. У гусаков и селезней имеется половой орган, который выпячивается при совокуплении, в спокойном состоянии он расположен в полости клоаки над прямой кишкой.
У самок хорошо развиты левый яичник и яйцевод, правый яичник и яйцевод редуцированы. Различают следующие отделы женских органов размножения: яичник, яйцевод, матку и влагалище, открывающиеся в клоаку (рис.45).
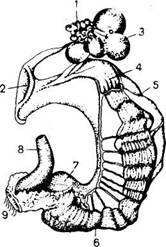
Рис. 45. Органы размножения курицы:
1 — яичник с фолликулами; 2 — воронка яйцевода; 3 — оболочка фолликула; 4 — белковая часть яйцевода; 5 — брыжейка; 6— перешеек; 7 — матка; 8 — толстая кишка; 9 — клоака
Яичник имеет форму продолговатой четырехугольной пластинки. Брюшной складкой он прикреплен к дорсальной стенке брюшной полости, а связкой - к яйцеводу, который представляет собой гроздевидный пакет отдельных яйцеклеток, находящихся в различной стадии развития (рис.46).
Яйцеклетка формируется на ранних стадиях эмбриогенеза птицы; ее развитие называется овогенезом, появляются овогонии и овоциты первого порядка; полностью процесс завершается после вывода цыпленка из яйца. У растущей птицы растет яйцеклетка и накапливается желток (вителлогенез). Способность к яйценоскости зависит от срока наступления половой зрелости, которая, в свою очередь, обусловлена видовыми и породными особенностями птицы.
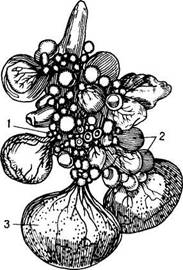
Рис. 46 . Внешний вид яичника курицы в период половой зрелости:
1 – первичный фолликул; 2 – растущие фолликулы; 3 – созревший фолликул
Кровоснабжение яичника осуществляется через яичную артерию, имеющую многочисленные разветвления, благодаря чему питательные вещества поступают сразу в несколько фолликул. Отток крови из яичника происходит через яичную вену. Каждый фолликул — это яйцеклетка, находящаяся на разной стадии формирования. Количество яйцеклеток (овоцитов, овогоний) у птицы отдельных видов может составлять несколько десятков тысяч, но созревают не все из них. При созревании фолликул увеличивается в объеме (у кур диаметр его равен 4 см). Значительное количество яйцеклеток достигает диаметра 1-2 см и претерпевает процесс атрезии (обратное развитие).
В результате истончения стенки фолликула происходит его разрыв и зрелая яйцеклетка попадает в расширенную часть яйцевода - воронку. Этот процесс называется овуляцией.
При заболевании, а также во время линьки птицы масса яичника уменьшается. Недоразвитие яичника и яйцевода отмечается при вирусном бронхите птицы, респираторных заболеваниях, в условиях неполноценного кормления.
Процесс формирования яйцеклетки и яйца регулируется нервно-эндокринной системой организма. Овуляция и выход яйцеклетки в воронку яйцевода происходит после снесения очередного яйца. В период формирования скорлупы увеличивается концентрация кальция в сыворотке крови птицы. Последовательность формирования яйца в яйцеводе приведена в табл.21.
Яйцевод представляет собой многослойную трубку. Если сделать поперечный разрез стенки яйцевода, то можно обнаружить многослойную структуру его. Покрыт он двумя слоями покровного эпителия, к которым прикреплены связки, подвешивающие яйцевод к позвоночному столбу. Яйцевод имеет вид спирали, с сильно развитыми гладкими мышцами. Отдельные мышечные волокна проникают в связки. Внутри яйцевод выстлан железистым эпителием, продуцирующим белок. В яйцеводе различают следующие отделы: воронку, белковую часть, перешеек, матку и влагалище.
Воронка — начальная часть яйцевода, где происходит оплодотворение яйцеклетки, покрыта цилиндрическим эпителием (у кур сильно расширена). Воронка яйцевода переходит в белковую часть (собственно отдел яйцевода), которая у хорошо несущихся кур собрана в складки, внутри выстлана цилиндрическим эпителием. Время нахождения яйца в белковой части яйцевода колеблется от 2 до 3 ч; вначале наслаивается плотный градинковый белок, затем остальные составляющие части яйца. У только что убитой курицы можно наблюдать перистальтику
стенки яйцевода, обеспечивающую медленное вращательное продвижение желтка, на который наслаиваются плотный и жидкий белок.
Перешеек расположен за белковой частью, выстлан изнутри кубическим эпителием, просвет яйцевода в этом участке сужается. В перешейке формируются подскорлупные оболочки яйца.
Матка - отрезок яйцевода, где происходит формирование скорлупы яйца. Стенки матки толстые и содержат большое
Вы здесь
Штеле А.Л., профессор кафедры интенсивных технологий в животноводстве, канд. с.-х. наук РГАУ — МСХА имени К.А. Тимирязева
Автор считает, что биологическую полноценность яиц и яичную продуктивность кур можно прогнозировать, начиная с развития яйцеклетки в процессе овогенеза.
Строение и состав птичьего яйца (ovo) соответствуют его природному назначению, связанному с размножением и воспроизводством потомства. Куриное яйцо представляет собой крупную яйцеклетку (ядро и цитоплазма), созревающую в естественной взаимосвязи с желтком. В полноценном яйце содержатся все питательные и биологически активные вещества, необходимые для развития эмбриона и вывода здорового молодняка, что определяет инкубационные качества и пищевую ценность яиц как натурального продукта питания.
Развитие и созревание яйцеклетки, образование и снесение яиц детерминированы геномом кур и проходят при взаимодействии с условиями среды: заданным световым режимом и оптимальным микроклиматом, нормированным кормлением . Полный процесс создания яиц в организме регулируется центральной нервной системой и находится под влиянием нейрогормональной деятельности организма. В наибольшей степени это связано с функциональной активностью гипоталамуса и гипофиза, гормонов яичника (эстрогены, прогестерон), что обеспечивает синхронность в созревании фолликулов, овуляции яйцеклетки-желтка, образовании и снесении яйца.
Биологическую полноценность яиц и яичную продуктивность кур можно прогнозировать, начиная с развития яйцеклетки в процессе овогенеза. Образование зрелой яйцеклетки (овогенез) проходит в три периода (стадии), продолжительность которых и продукты, сведены нами в таблицу 1.
Таблица 1. Периоды и стадии овогенеза у высокопродуктивных кур (по обобщенным данным)
Первый период овогенеза отмечается в эмбриогенезе кур, когда первичные зародышевые клетки (овогонии) обособляются на 8-11-е сутки развития эмбриона. Эти клетки многократно делятся путем митоза и к суточному возрасту преобразуются в первичные яйцеклетки (овоциты) с диплоидным набором хромосом. Вокруг каждой из них (около 4 тыс.) формируется фолликул, благодаря которому осуществляется взаимосвязь между развивающимся овоцитом и яичником.
Во втором периоде овогенеза яйцеклетки растут и развиваются за счет цитоплазмы и накопления желточного материала. В первой стадии (0-6 нед.) отмечается увеличение ядра и цитоплазмы, формируется зародышевый диск, а диаметр яйцеклетки достигает 0,5-1,0 мм. Вторая стадия овогенеза длится до 16-недельного возраста курочек и характерна накоплением желтка в фолликулах по мере роста и развития яичника и яйцевода.
В процессе овогенеза питательные и биологически активные вещества интенсивно синтезируются и/или концентрируются в печени и с током крови поступают в фолликулы. Функционирование системы овоцит-фолликул обеспечивает биосинтез в желтке РНК и стероидов, стимулирующих созревание самой яйцеклетки. При этом гипофиз активно вырабатывает фолликулостимулирующий гормон, который ускоряет рост и развитие овоцитов в фолликулах.
В третьем периоде овогенеза, в 16-17-недельном возрасте курочек, происходит созревание яйцеклетки. При этом ускоренное накопление желтка последовательно отмечается в 5-6 больших фолликулах яичника и занимает около 1 нед. Непосредственно перед овуляцией яйцеклетка представляет собой одну клетку, заполненную питательным веществом (желток), окруженную плотной вителлиновой оболочкой, а ее ядро расположено в бластодиске.
Первое деление мейоза в ядре яйцеклетки проходит в яичнике до начала овуляции. После перемещения яйцеклетки-желтка в воронку яйцевода отмечается второе деление мейоза, и образуется половая гамета с гаплоидным набором хромосом, способная к оплодотворению. При слиянии яйцеклетки со сперматозоидом на стадии зиготы восстанавливается диплоидный набор хромосом.
В репродуктивный период каждая яйцеклетка совершает длительный путь роста и развития до созревания и овуляции (17-18 нед.) независимо от того, будет ли она оплодотворена в яйцеводе или нет. По мере продвижения желтка по яйцеводу образуются белок и скорлупа, что завершается снесением полноценного яйца. В яйцеводе яйцо формируется обычно за 23-26 ч, в том числе в воронке - 20 мин, белковом отделе - 3 ч, перешейке - 1 ч, матке - 16-20 ч (в среднем - 18-19 ч).
Образование полноценных яиц
Яичная продуктивность кур (яйценоскость х масса яиц) определяется физиологическим состоянием организма птицы и интенсивностью обмена веществ. Сложный и длительный процесс роста и развития яйцеклетки-желтка в яичнике (0-17 нед.) является ограничительным фактором генома кур при формировании продуктивности в процессе онтогенеза. Фенотипические изменения признаков при давлении селекции расширяют физиологические границы организма за счет интенсивного функционирования яичника/яйцевода и быстрого метаболизма в организме. Известный факт снесения курицей породы леггорн 361 яйца за 364 дня практически при 100%-ной яйценоскости можно считать пределом яичной продуктивности отдельной особи.
Высокопродуктивные куры-несушки яичных кроссов отличаются скорым накоплением питательных и биологически активных веществ в последовательно созревающих 4-5 крупных фолликулах. В экспериментах установлено, что максимальный темп переноса питательных веществ из плазмы крови в фолликул в расчете на единицу его поверхности отмечается при массе овоцита 2,5 г.
Можно считать, что это начальный фолликул в любой серии нескольких быстро развивающихся фолликулов в яичнике. Затем, с увеличением поверхности каждого последующего до овуляции фолликула, скорость отложения желточного материала нарастает. В самом крупном из них масса желтка в наибольшей степени повышается за 24 ч до овуляции, а его диаметр достигает 35 мм.
При относительно невысокой яйценоскости, характерной для яичных пород (кроме белых леггорнов), мясо-яичных пород и линий, в яичнике одновременно формируются 7-10 крупных фолликулов При таком числе развивающихся фолликул, особенно во второй половине продуктивного периода, чаще отмечается десинхронизация овуляции и яйцекладки. Это является одной из причин образования двухжелтковых яиц с большой массой - 75-80 г и более. Отмечено, что яйца с двумя желтками откладывают и молодые куры, чей цикл яйцекладки и ритм яйценоскости еще не синхронизированы.
Немаловажным фактором высокой и устойчивой продуктивности является биологический цикл яйцекладки, характерный для каждой особи. По данным исследователей, при яйценоскости кур до 300 яиц цикл непрерывной яйцекладки составляет 30-50 яиц с интервалом (паузой) в среднем 2 дня. При напряженной яйценоскости кур выше 300 яиц отмечен более длинный цикл - 40-80 яиц, также с небольшим интервалом, что определяет ритм яйценоскости. При этом среднегодовая продуктивность кур за 52 нед. может составить 330-340, а на начальную несушку - 310-320 яиц при сохранности 94-95%.
Таким образом, общее время образования биологически полноценного яйца, способного к оплодотворению, в среднем составляет 6-7 суток. Можно считать, что чем меньше крупных фолликулов (4-5) в яичнике, тем быстрее происходит накопление желтка в них и созревание яйцеклетки. Ранее в яичнике продуктивных кур отмечалось 7-8 и более больших фолликулов, число которых определяло яйценоскость птицы.
Периодизация выращивания молодняка
Формирование рспродѵктивных органов (яичник и яйцевод), развитие и созревания яйцеклеток, половая скороспелость при выращивании ремонтных курочек сопряжены с периодами их роста и развития и, соответственно, с фазами кормления. За последние 70 с лишним лет интенсивной селекции яичных кроссов и гибридизации отмечено значительное снижение возраста снесения первого яйца. Срок половой скороспелости курочек за это время сократился в 1,5 раза: с 6 мес. (26-27 нед.) до 4 мес. (17-18 нед.), что привело к аналогичному сокращению периода постэмбрионального развития в онтогенезе. Основные показатели ограничительных факторов при определенном соотношении овогенез/онтогенез приведены в таблице 2. Существенно более быстрый рост и развитие курочек для современных яичных кроссов, контролируемый по состоянию костной ткани, привел к слабости костей ног и крыльев, отмечаемой в начале продуктивного периода. Эта проблема решается благодаря нормированному кормлению при выращивании молодняка и строгому ограничению живой массы несушек к началу яйцекладки (1,2-1,3 кг).
Таблица 2. Контролируемые биологические и зоотехнические факторы при выращивании курочек (на примере кросса Хай-Лайн W 98)
Яичная продуктивность кур
В практике и племенного, и промышленного птицеводства принято использовать кур в течение 52 нед. продуктивного периода, или 70 нед. жизни. Оценка яйценоскости на начальное поголовье кур позволяет контролировать сохранность птицы. Для современных яичных кроссов характерны сходные показатели яичной продуктивности кур. У белых и коричневых молодых курочек возраст половой скороспелости и достижения 50%-ной яйценоскости практически одинаков, так же как яйценоскость и масса яиц.
В ряде исследований показано, что физиологические нормы овуляций при наивысшей продуктивности кур (на уровне 90% и более) близки к биологическому пределу. Поэтому большое внимание уделяется селекции: на повышение жизнеспособности и устойчивости к заболеваниям, состоянию оперения кур. Селекция яичных кроссов по продуктивности позволила увеличить яйценоскость до 350-360 яиц на начальную несушку за 62 нед. продуктивного периода (табл. 3).
Таблица 3. Яичная продуктивность яичных кур белых и коричневых кроссов
Выделено девять основных факторов формирования продуктивности, из них пять биологических: геном вида; давление селекции; овогенез - образование зрелой яйцеклетки, способной к оплодотворению; эмбриональный и продуктивный периоды. В числе основных зоотехнических факторов - заданный световой режим, оптимальный микроклимат, нормированное ограниченное кормление, технология производства яиц.
Отметим, что постоянное давление селекции на увеличение яичной продуктивности сопряжено и поддерживается снижением живой массы птицы. В определенной мере эти показатели нивелируются в селекционных программах компаний (и фирм), а в племенных хозяйствах ограничивают яйценоскость и массу яиц прародительских и родительских линий. Это позволяет поддерживать высокую яичную продуктивность при скрещивании линий у гибридных кур промышленного стада. Сегодня формирование яичной продуктивности кур во многом определяется строго контролируемой живой массой. От суточного возраста до завершения продуктивного периода заданная живая масса стала главным показателем роста, развития и физиологического состояния организма птицы.
Взаимодействие биологических факторов и зоотехнических нормативов в условиях промышленного птицеводства позволяет поддерживать среднегодовую (52 нед.) интенсивность яйценоскости несушек на уровне 82-85%, а при пике яйцекладки - 90-95%. При этом вся жизнедеятельность птицы от светонепроницаемых птичников до завершения продуктивного периода выстроена в рамках «роботизации» живого организма. Длительное время напряженной (стрессы), непрерывной яйценоскости кур-несушек (без линьки) в несвойственных им условиях содержания определяет необходимость использовать отдельные элементы «технологии благополучия», реализуемой в большинстве европейских стран.
Биологическая полноценность яиц
Куриное яйцо как биологический объект и натуральный продукт питания имеет присущие ему морфологические признаки (строение), физико-химические свойства и биохимический состав. Биологически полноценное яйцо включает зрелую яйцеклетку (половую гамету) на поверхности желтка и многослойный белок в прочной скорлупе. При сбалансированном кормлении кур в полноценном яйце содержатся белки (незаменимые аминокислоты), липиды и жирные кислоты, незначительно (около 1%) углеводы, витамины и микроэлементы, другие биологически активные вещества.
При содержании кур в племенных хозяйствах с петухами оплодотворенные яйца передают на инкубацию, а биологическая ценность определяется биохимическим составом, оплодотворенностью и выводимостью яиц. В промышленных стадах от кур-несушек получают неоплодотворенные яйца, которые также относятся к биологически полноценным продуктам. Полноценность пищевых яиц определяется не только организма человека в питательных веществах и энергии (калорийность).
Таблица 4. Химический состав, калорийность и масса составных частей яиц
Примечание: * - с 2000 года при пересчете количества яиц (шт.) в весовые показатели (кг, т) средняя масса яиц принята 60 г.
В таблице 4 приведены химический состав и калорийность яиц, пересчитанные на среднюю массу 60 г. Особенно богат питательными веществами желток яиц, что определяет влажность оптимального соотношения составных частей яйца, которое во многом обусловливается его массой. В среднем белок, желток и скорлупа куриного яйца имеют пропорцию 6:3:1 или 59-61, 29-31 и 10-11 %. Размах колебаний этих показателей соответственно - 53-69, 24-36 и 8-14 % определяется генотипом (породой, кроссом), возрастом несушек и другими факторами, влияющими на массу яиц.
Содержимое яйца (желток, многослойный белок) отличается от всех других продуктов питания высокой полноценностью белков. Считается, что незаменимые аминокислоты - триптофан, метионин, лизин «в идеальном пищевом белке» должны иметь соотношение 1,0:3,5:5,5. Наиболее близким к этому показателю среди животных продуктов является белок куриного яйца — 1,6:3,3:6,9.
Количество и соотношение незаменимых аминокислот, их соответствие потребностям человека характеризуют биологическую ценность белков яиц.
Заключение
Созревание яйцеклетки в естественной взаимосвязи с желтком в фолликулах яичника, образование белка и скорлупы в яйцеводе, биологический цикл яйцекладки и ритм яйценоскости совершенно синхронизированы у высокопродуктивных кур. Интенсивное накопление желтка в нескольких быстрорастущих фолликулах яичника и возможная ежедневная овуляция при длительных циклах являются границей яичной продуктивности кур.
Чем меньше крупных фолликул (5-6 не более), одновременно находящихся в яичнике на стадии созревания яйцеклетки-желтка, тем выше продуктивность кур-несушек. Количество фолликулов в яичнике на этой стадии является одним из показателей физиологического состояния органов размножения и потенциала высокой яйценоскости кур.
В промышленном птицеводстве кур-несушек используют в течение первого продуктивного периода (52 нед.) при высокой интенсивности яйценоскости (82-85%), что обеспечивает эффективность производства пищевых яиц. Для некоторых яичных кроссов срок продуктивного использования кур может составлять 62 нед. и более.
Белок куриного яйца при оптимальном для организма человека количестве и соотношении незаменимых аминокислот принят за эталон биологической ценности. Иными словами, аминокислотный состав любого животного или растительного белка оценивают по отношению к незаменимым аминокислотам яичного белка.
Переход в промышленном птицеводстве на более продолжительное использование кур-несушек (14-15 мес.) при строго контролируемой живой массе, ограниченном кормлении и интенсивной технологии предопределяет новые подходы к направленному выращиванию ремонтного молодняка и фазовому кормлению кур-несушек, а также применение отдельных элементов технологии благополучия птицы.
Штеле А.Л., профессор кафедры интенсивных технологий в животноводстве, канд. с.-х. наук РГАУ — МСХА имени К.А. Тимирязева
журнал "Птица и птицепродукты" №6, 2011 г
Читайте также: