
Наследственные заболевания у лошадей
Опубликовано: 30.04.2024
Автор: В. ПАРШИКОВА
Номер журнала: ЗМ №4(12)1999
Наследственные болезни лошадей до последнего момента практически не были изучены и оставались «тайной за семью печатями». Однако люди давно подметили, что некоторые заболевания являются врожденными, либо возникают в определенном возрасте, как бы без видимых причин, но в то же время повторяются у лошадей, имеющих общее происхождение. Со временем исследователи и коневоды пришли к мысли о передаче ряда болезней или дефектов по наследству, но достоверных научных данных об их природе не было вплоть до середины нашего столетия.
И даже изобретения микроскопа не расставило все точки над и i, хотя и позволило установить, что наследственные признаки передаются из поколения в поколение через сперматозоиды и яйцеклетки, но каким образом происходит этот сложный механизм обмена информацией, известно не было.
Первый действительно научный шаг вперед сделал Грегор Мендель, опубликовав научную статью, заложившую основу современной генетики. И сейчас, когда человечество накопило достаточный багаж знаний, можно с уверенностью сказать, что за генетическими исследованиями стоит наше будущее.
В этой статье мы попробуем вкратце объяснить причину возникновения генетических заболеваний у лошадей, но для более полного понимания нам придется сделать маленький экскурс в колыбель всех биологических наук — цитологию.
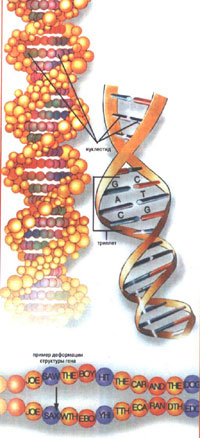
Итак, каждая лошадь состоит из многих триллионов клеток, различных по размеру, форме и функциям. Внутри ядра каждой клетки находится 64 х-образные хромосомы, соединенные в 32 пары (к примеру, у человека 46 хромосом, соединенных в 23 пары.) Именно они содержат полную генетическую инструкцию для построения и фунциклирования каждой клетки и, следовательно, всего организма в целом. Хромосомы состоят из двойной спирали ДНК (дезоксирибонуклеиновая кислота). Ген — это функциональный сегмент ДНК, ответственный за каждую характеристику лошади, от цвета шерсти до работы ее пищеварительного тракта. Геном лошади содержит более 700 тысяч генов.
Молекулярная модель ДНК огромная по молекулярным масштабам спираль состоящая из двух закрученых нитей, на которых подобно бусам "насажены" 4 типа нуклеотидов (Аденин-А, Тимин-Т, Гуанин-Г и Цитозин-Ц), идущих в строго определенном порядке, и образующих между собой пары; Аденин может образовывать пары, только с Тимином, а Гуанин только с Цитозином. У лошадей в геноме примерно 3 биллиона таких пар. Зачем все так сложно? А затем, что в построенной таким образом спирали каждая нить содержит сведения о своей «напарнице», и, таким образом, зная содержимое одной нити всегда можно достроить ей парную. Это важнейшее условие для воспроизводства всего живого!
Для того чтобы не утомлять вас сложной терминологией, представьте себе, что сам генетический код — это огромная книга, а ген будет в ней только предложением, но некоторые гены в этой книге столь длинны, что занимают несколько страниц. Гены в свою очередь состоят из слов — триплетов, а каждый триплет состоит из трех нуклеотидов — т. е. букв.
В силу необъяснимых естественных причин, или под влиянием искусственных факторов (самый распространенный из них радиация) начинает деформироваться структура самого гена на молекулярном уровне — происходит мутация, которая, в свою очередь, изменяет программу гена. То есть дефектный ген (подвергшийся мутации) еще на ранних стадиях развития зародыша лошади "дает не-правильную инструкцию" для построения "подотчетных" ему клеток. И хорошо, если окажется, что действие данной мутации будет для организма нейтральной и не опасной для жизни, но зачастую мутации приводят к выкидышу эмбриона на ранних стадиях, или проявляются позже в форме тех или иных генетических заболеваний.
Есть еще одна немаловажная проблема, встающая на пути генетического исследования лошадей; дело в том, что само по себе наличие у животного дефектного гена не всегда вызывает клиническое проявление болезни: большинство таких генов — рецессивны (более слабо влияющие на формирование тех или иных признаков и подавляемые сильными генами — доминатными). Это означает, что лошадь может заболеть наследственной болезнью лишь тогда, когда оба ее родителя будут иметь дефектный ген, а может и не заболеть, а стать носителем деструктивного гена, и, в свою очередь, превратиться в так называемую «мину замедленного действия». В итоге один единственный носитель какого-нибудь дефекта (например, жеребец-производитель) может передать его сотням своим потомков в нескольких поколениях, и его дети будут благополучно использоваться в селекционной работе, пока у одного из них не совпадут два рецессивных гена и болезнь не проявиться в полную силу. Само собой разумеется, что владельцы ценных племенных лошадей откажутся признать у своих питомцев наличие рокового генетического дефекта до предъявления им веских научных подтверждений. А пока будут тянуться всевозможные дрязги между ветеринарами и владельцами, и ученые будут ставить эксперименты для научного обоснования — пройдут годы, и генетические мутации станут частью генетического наследия той или иной породы или линии. В этом и заключается опасность и непредсказуемость генетических болезней.
Как же можно предотвратить неприятные сюрпризы природы? К сожалению, весь аспект этого вопроса упирается в одно — тщательное исследование генетического материала на молекулярном уровне, требует колоссальных финансовых затрат. Но исследователи не останавливаются и продолжают свои поиски, и, к счастью, продвигаются вперед. Например, изобретен так называемый рабочий словарь для хромосомного лексикона лошади. С таким словарем— известным как генетическая карта — ученые
могут разработать точные тесты для идентификации типичных специфических мутаций. Одним словом научная база разведения лошадей стоит на пороге коренных перемен.
В Америке, где испытаниям чистокровных лошадей и рысаков уделяется особое внимание, ученые всерьез озабочены поискам «гена скорости». Как. считают исследователи, это открытие поможет им получить 100% элитных лошадей с непревзойденными беговыми или скаковыми качествами. А если учесть, что призовые на крупных призах переваливают за миллион долларов, то рвение ученых становится понятным.
Подобное предположение не столь невероятно, как кажется: аналогичная технология уже с у спехом применяется генными инженерами для получения более продуктивных линий молочных коров.
Дискуссия о «гене скорости» стала основной для некоторых исследований крупнейших американских научных центров и лабораторий. В результате ученые пришли к выводу, что за «скорость» отвечают сразу несколько компонентов разных генов.

Генетика: от рытья нор до учебы в колледже
18 июня 2013 года. ТрВ № 131, c. 14, "Исследования"
Михаил Гельфанд
Рубрика: Исследования
В мае были опубликованы результаты генетического анализа более 125 тысяч человек, обнаружившего три полиморфизма (участка в геноме, в котором существуют различные варианты), которые коррелируют с образовательными достижениями [1]. Лица с благоприятными вариантами за свою жизнь проводят в учебных учреждениях в среднем на один месяц дольше и с чуть большей вероятностью заканчивают колледж (ясно, впрочем, что это скоррелированные параметры). Учет каждого из трех генетических факторов объясняет примерно 0,02% вариаций в образовательном уровне; комбинация всех вариантов (включая статистически незначимые) объясняет 2% разнообразия. Казалось бы, это не выглядит впечатляюще — но не надо недооценивать сложности исследуемого признака, да и невозможность произвести прямые эксперименты на объекте исследования.
pinterest.com
Лошади тоже не очень удобный объект для экспериментов, но зато хороший для генетических исследований благодаря тщательным записям родословных и существованию чистых пород. В прошлом году группа шведских, исландских и американских исследователей установила ген, определяющий иноходцев [2]. Работа началась с исследования полногеномных ассоциаций у 70 исландских лошадей, 40 из которых были способны скакать иноходью. Был обнаружен единственный полиморфизм, сильно ассоциированный с этой способностью. Секвенирование (определение последовательности) области генома вокруг этого полиморфизма у 352 лошадей показало,что все иноходцы гомозиготны по нонсенс-мутации гена DMRT3 — у них этот ген вообще не работает, с него не читается полноценный белок. У неиноходцев такая ситуация наблюдалась в 31% случаев. На самом деле среди иноходцев было единственное исключение. Но, как сказано в статье, тщательное изучение дневников соревнований показало, что эта лошадь, скорее всего, была ошибочно отнесена к иноходцам. То же, что среди лошадей с мутацией много неиноходцев, неудивительно: это просто отражает особенности тренировки.
Частота мутации в различных породах сильно варьируется: у пород, которые отбирались на способность удерживать заданный аллюр, не сбиваясь на большой скорости на галоп, эта мутация часта, а у пород, отобранных на выездку или на скорость, она не встречается. Еще одно генетическое исследование было проведено на шведских скакунах (Swedish Standardbred): эта порода была выведена смешением американских скакунов (American Standardbred, в этой породе мутация зафиксировалась) и французских рысаков (French Trotter, в основном имеющих обычный вариант). Тем самым, мутация у шведской породы имеет промежуточную частоту, что дало возможность исследовать ее распределение в зависимости от спортивных результатов лошадей. Оказалось, что скакуны, гомозиготные по мутации, выиграли в среднем в 3 раза больше призовых денег, чем гетерозиготы.
wikimedia.org
Но изучать молекулярно-генетические механизмы на лошадях неудобно, и дальнейшее исследование было проведено на обычных лабораторных мышах, которых помещали на стандартную установку для изучения движений. Оказалось, что мутантные мыши испытывают сложности с бегом на больших скоростях, у них увеличена длина шага, а у новорожденных мышат повышена относительная частота некоординированных движений конечностей. Эксперименты на молекулярном уровне показали, что клетки, в которых работает этот ген,- это ингибирующие интернейроны спинного мозга, непосредственно связанные с моторными нейронами и осуществляющие координацию движений левых и правых конечностей.
Продолжим разговор о мышах, точнее об американских оленьих мышах Peromyscus, отдаленно родственных обычным мышам из рода Mus. Peromyscus — самый многочисленный род млекопитающих. Два недавно разошедшихся вида, P. polionotus и P. maniculatus, роют норы разной планировки: P. maniculatus, как и другие представители рода, роет простые неглубокие норы, а P. polion

Ежегодный информационно-аналитический альманах
Ахал-Теке информ за 2006 год
"Наука и практика"
"Ставропольский сфинкс"
В октябре 1982 года в составе экспертной комиссии, проводившей выставку - выводкуахалтекинский лошадей на Ашхабадском ипподроме, я поснтила конный завод "Комсомол". И там увидела редчайшую картину: По внутренниму двору конной части следом за начконом Аширом Аннаевым, держащим в руке ведро с овсом, шла соловая кобыла, а за ней два жеребенка. Неужели двойня? Один жеребенок, хотя прыгал и резвился так же, как и его братец, выглядел как то странно, и я не сразу поняла, в чем эта странность: в серо - розовой пятнистой масти, в очень длинных ушах и необычайно тонком хвосте?
- Это голяк. - сказала Мария Даниловна Черкезова. - Он родился без волосяного покрова и долго не проживет. До первых холодов.
Этот давний случай я сразу вспомнила в сентябре 2005 года в Ставропольском конном заводе, куда традиционно приглашались участники и гости скакового митинга. И снова пришла в изумление, издали не разобрав, кто это, похожий на огромного серого дога с розовыми ногами, гуляет на лужайке? Хвост, самая отличительная лошадиная часть, был гол и походил на хвост дога, а в остальном перед нами оказалась прелестная, упитанная кобылка. Она могла бы считаться воронной масти, если б обладала ею в нужной степени, с белыми отметинами на ногах, и эти отметины отливали розовым цветом, как и проточинка на голове. А на мордочке, мягкой и нежной на ощупь, мимические морщинки. Кто-то тут же окрестил кобылку "сфинксом", а кто - то добавил - "ставропольский сфинкс". Это был классический голяк, без единой волосинки на теле.
Кобылка родилась в 2004 году от Джейнара и Туйчи и ее, как неперспективную для жизни в общих конюшенных условиях, отдали на попечение работнику кончасти Владимиру Вискалину. Вся семья участвовала в заботливом уходе за голой кобылкой: шили теплые попоны, оберегали от сковняков, старались повкуснее накормить. В воспитании помогла домашняя собака, с которой Малышка, так назвали кобылку, крепко подружилась и даже переняля некоторые особенности поведения. Малышка четко выполняет комынды "Ко мне!", "Рядом!", "Стоять!". Пятилетняя дочка Вискалиных смело восседает на ней без седла, недоуздок связали и надели исключительно для важности.
Итак, "ставропольский сфинкс" доказал, что в заботливых руках вполне жилец. Что дальше?
Ежегодный информационно-аналитический альманах
Ахал-Теке информ за 2006 год
"Наука и практика"
"Ставропольский сфинкс"
В октябре 1982 года в составе экспертной комиссии, проводившей выставку - выводкуахалтекинский лошадей на Ашхабадском ипподроме, я поснтила конный завод "Комсомол". И там увидела редчайшую картину: По внутренниму двору конной части следом за начконом Аширом Аннаевым, держащим в руке ведро с овсом, шла соловая кобыла, а за ней два жеребенка. Неужели двойня? Один жеребенок, хотя прыгал и резвился так же, как и его братец, выглядел как то странно, и я не сразу поняла, в чем эта странность: в серо - розовой пятнистой масти, в очень длинных ушах и необычайно тонком хвосте?
- Это голяк. - сказала Мария Даниловна Черкезова. - Он родился без волосяного покрова и долго не проживет. До первых холодов.
Этот давний случай я сразу вспомнила в сентябре 2005 года в Ставропольском конном заводе, куда традиционно приглашались участники и гости скакового митинга. И снова пришла в изумление, издали не разобрав, кто это, похожий на огромного серого дога с розовыми ногами, гуляет на лужайке? Хвост, самая отличительная лошадиная часть, был гол и походил на хвост дога, а в остальном перед нами оказалась прелестная, упитанная кобылка. Она могла бы считаться воронной масти, если б обладала ею в нужной степени, с белыми отметинами на ногах, и эти отметины отливали розовым цветом, как и проточинка на голове. А на мордочке, мягкой и нежной на ощупь, мимические морщинки. Кто-то тут же окрестил кобылку "сфинксом", а кто - то добавил - "ставропольский сфинкс". Это был классический голяк, без единой волосинки на теле.
Кобылка родилась в 2004 году от Джейнара и Туйчи и ее, как неперспективную для жизни в общих конюшенных условиях, отдали на попечение работнику кончасти Владимиру Вискалину. Вся семья участвовала в заботливом уходе за голой кобылкой: шили теплые попоны, оберегали от сковняков, старались повкуснее накормить. В воспитании помогла домашняя собака, с которой Малышка, так назвали кобылку, крепко подружилась и даже переняля некоторые особенности поведения. Малышка четко выполняет комынды "Ко мне!", "Рядом!", "Стоять!". Пятилетняя дочка Вискалиных смело восседает на ней без седла, недоуздок связали и надели исключительно для важности.
Итак, "ставропольский сфинкс" доказал, что в заботливых руках вполне жилец. Что дальше?
А дальше попробуем разобраться в этом явлении по науке.
Мы просмотрели все восемь томов ГПК, в которые внесены данные на 2994 матки с их приплодом, и обнаружили официально записанными голяками 9 голов, причем все они пали. К ним прибавим еще двух, описанных О. Карауш в 1938 году, до выхода первой племенной книги и еще двух, обнаруженных нами, но не значащихся в ГПК голяками. Это тот самый Кимбилер, увиденный воочию в конном заводе "Комсомол" в 1982 году, и кобылка Нагая, 2001 года рождения, вычисленная дедуктивным путем (судя по имени, а так же по ее матери Мариуле, замеченной в "деле" голяков). Так же прибавим еще двух, выясненных путем опроса: не записанного в регистр и павшего вскоре после рождения в 2002 году солового жеребчика и, прожившую шесть месяцев, вороную кобылку рождения 2005 года от Машука и Мимозы, названную Мумией. мумия родилась довольно крупной и очень породной, однако через месяц стала отставать в росте от сверстников. Она часто стадала насморком и расстройством пищеварения, умерла от простуды. И наконец, нашу героиню, записанную в регистре молодняка без клички. И того получилось 16 голяков. Как мы заметили, не всех голяков показывают. Они могут скрываться под "мертво и слаборожденными" и просто под записью "пал" в списке плодовой деятельности кобылы. Но даже если среди них и окажутся голяки, то общее количество их врят ли превысило бы вдвое нашу цифру, то есть 32. От 2994 маток это максимально сосавило бы 1;2%, а если отнести к общем массе полученного от этих кобыл потомства (возьмем в сред
А дальше попробуем разобраться в этом явлении по науке.
Мы просмотрели все восемь томов ГПК, в которые внесены данные на 2994 матки с их приплодом, и обнаружили официально записанными голяками 9 голов, причем все они пали. К ним прибавим еще двух, описанных О. Карауш в 1938 году, до выхода первой племенной книги и еще двух, обнаруженных нами, но не значащихся в ГПК голяками. Это тот самый Кимбилер, увиденный воочию в конном заводе "Комсомол" в 1982 году, и кобылка Нагая, 2001 года рождения, вычисленная дедуктивным путем (судя по имени, а так же по ее матери Мариуле, замеченной в "деле" голяков). Так же прибавим еще двух, выясненных путем опроса: не записанного в регистр и павшего вскоре после рождения в 2002 году солового жеребчика и, прожившую шесть месяцев, вороную кобылку рождения 2005 года от Машука и Мимозы, названную Мумией. мумия родилась довольно крупной и очень породной, однако через месяц стала отставать в росте от сверстников. Она часто стадала насморком и расстройством пищеварения, умерла от простуды. И наконец, нашу героиню, записанную в регистре молодняка без клички. И того получилось 16 голяков. Как мы заметили, не всех голяков показывают. Они могут скрываться под "мертво и слаборожденными" и просто под записью "пал" в списке плодовой деятельности кобылы. Но даже если среди них и окажутся голяки, то общее количество их врят ли превысило бы вдвое нашу цифру, то есть 32. От 2994 маток это максимально сосавило бы 1;2%, а если отнести к общем массе полученного от этих кобыл потомства (возьмем в среднем по 7 жеребят от каждой) - то и вовсе 0,15%. Какова динамика появления голых жеребят? После 1938 года первый голяк был зарегестрирован только в 1960 году, и их количество неравномерно по десятилетиям: 60-е годы - 4 головы, 70-е, 80-е - 3, 90-е - 2, а за пять лет первого десятилетия двухтысячных - 5 голов. Увеличение числа голяков, вероятно, связанно с общим увеличением поголовья, а отстутствие в определенные периоды - стремление скрыть это явление. 8 голов родилось в Туркмении (причем мы не имеем точных сведений за последние 10 лет), 7 - в России, 1 - в Германии, что вполне укладывается в числовой ряд поголовья в этих странах.
Первое и единственное объясние в отечественной литературе феномена голых жеребят мы находим в журнале "Коневодство", 1938г. № 8-9 в статье доцента Туркменского сельскохозяйственного института им. М. И. Калинина О. Н. Карауш "голые жеребята". Ольга Николаевна описывает виденных ею двух голых жеребят, родившихся в марте 1938 года в колхозе им. Ворошилова. Первый жеребенок прожил 2 месяца и умер от сильно поноса, хотя всю свою коротенькую жизнь выглядел вполне здоровым. Этот жеребенок родился совершенно безволосым и не имел молочных резцов (не аномалия), встал на ноги самостоятельно и начал сосать мать. В 2-х месячном возрасте имел две пары молочных резцов (зацепы и стредние), был крупным, правильно сложенным, типичным ахалтекинцем. Кожа, образовавшая мелкие складки, была темно-серой (слоновой) окраски, а отметины - розового цвета (как и у нашей Малышки). Под лупой рассматривались отдельные, редкие волоски на спине, крупе, лопатках, на конце хвоста, спереди на запястных и сзади на скакательных суставах. Также установлено наличие потоотделения. Второй голый жеребенок был также нормально развитый, самостоятельно встал на ноги и сосал мать, но прожил лишь 3 дня, умер вероятно от простуды, така как стояла холодная погода. Версию о недоношенности принять нельзя. Причину, по мнению Ольги Николаевны, следует искать в происхождении жеребчиков, они близкие родственники. У них общий отец, гнедой Дор Дели 1930 г. р. от Топорбая и Бек Назар Бай. Мать первого - буланая Меле-Хан 1929г. (Меле Куш - ах. коб.), (В ГПК она значится 301 Меле неизвестного происхождения). Матерью второго голяка является дочь Меле-Хан - буланая Мумме (Ай) от Мамеда, сына Топорбая. Анализируя родословные голых жеребят, О. Н. Карауш видит причину их безволосости в наличии инбридинга на Топорбая (III - II) у второго жеребенка. Что же касается инбридинга на мат
Что же касается инбридинга на мать Меле Куша - Ораз Нияз Карадашли (III-IV у первого жеребенка и IV-V-IV - у второго), отмеченного О. Н., иы будучи уверены в происхождении кобылы Меле-Хан, не можем принять его во внимание. Однако ставим, как основную, версию о влиянии инбридинга на проявление признака бесшерстности.
Проанализировав родословные наших "героев", мы обнаружили наиболее подозрительных жеребцов, могущих считаться носителями признака бесшерстности. Самым ранним подозреваемым мы считаем 87 Дор Депеля, несущего в себе кровь 244 Топорбая и 44 Бек Назар Дора, которые сместе со своими отцами и братьями, так же входят в этот ряд. В неменьшей степени подозреваются 213 Сапар Хан, 220 Случай (внук по матери 44 Бек Назар Дора), 448 Кир Сакар. Уверенность в этом подкрепляют потомки названных жеребцов - "рекордисты" Жеребец 796 Сере (Сайван - Еркуль) - внук Сапар Хана и правнук Бек Назар Ала (отца Бек Назар Дора). От него получено два голяка, причем в классическом сочетании с внучками Сапар Хана. Кобыла 1223 Каракеик (Сухты - Ша Гельды) - внучка Сапар Хана по отцу и Бек Назар - по матери. От нее так же полученно два голяка: от 796 Сере и от 736 Кеймира (в обоих случаях - имбридинг на Сапар Хана (III-III). Каракеик известна так же как мать двух знаменитых Мелекушей, подаренных в свое время в Англию и Индию. Но они родились от Меллека, линии Скака, и с шерстным покровом, у них было все в порядке. жеребец 736 Кеймир (Кир Сакар-Синица) - внук Сапар Хана по матери. От него имелось так же два "голыша": от упомянутой Каракеик и от 1109 Алкеик, внучки Сапар Хана. Жеребец 1054 Гылкуюк (721 Каплан - 1555 Келята) - внук 736 Кеймира. У него так же два голяка: от 2206 Гортензии (170 Санджар - 1934 Гульсар) и от самой 1936 Гульсар (900 Менджу - 1478 Гырды). В родословной Гульсар встречаются все подозреваемые: Кеймир, Кир Сакар, Сапар Хан, Случай, Бек Назар Ал. Так же прославились в этом деле 2001 Мариула (919 Сере - 1614 Мерджен), несущая в себе кровь Сапар Хана, Бек Назар Ала, Случая. От 943 Арслана (697 Гелишикли - 1112 Алсона), внука Сапар Хана, несущего так же кровь Бек Назар Ала, она родила ту самую кобылку по имени Нагая. Дочь Мариулы - 2860 Мрия от 1201 Кавказа - также родила голенького. А дочь тех же Мрии и Кавказа - 2818 Мекка проходит в "деле" недолго прожившей Мумии - она ее прабабушка по отцу. "Помог" бедной Мумии и имбридинг на 2001 Мариулу (III-IV).
Итак, совершенно очевидно, что бесшерстность - наследственный признак. Причина кроется не в имбридинге (не секрет, в ахалтекинской породе достаточно особей инбредного происхождения, но с нормальным шерстным покровом), а в типе наследования этого признака, который нам предстоит разгадать. Хотя у наших голяков, как правило. наличествует инбридинг на одного или нескольких производителей, но никогда не тесный, не кровосмешение.
При внимательном подборе, зная виновников в лицо, можно, как исключить, так и провоцировать появление голых лошадей. Вопрос лишь в том, хотим ли мы этого? А почему бы нет? К прочим уникальным качествам ахалтекинской породы можно добавить еще одно. Ведь в других породах: арабской, английской скаковой, орловской рысистой (мы просмотрели ГПК) - такого признака не обнаружено. (Кстати О. Н. Карауш упоминала об афганских голых лошадях, наши поиски этих лошадей не увенчались успехом)
Признак бесшерстности встречается и у крупного рогатого скота, птиц, кошек, собак и даже людей. У собак, кошек, кур бесшерстность закреплена и стойко передается потомкам, что позволило выделить отдельные породы. Голые животные, в частности кошки, имеют свои особенности развития: Они рождаются более крупными и развитыми, но позже отстают в росте от сверстников кроткошерстных пород (это успели отметить и у голых жеребят). Половое созревание у них наступает позже, и сервис-период длится вдвое дольше. Также замечены и некоторые различия по биохимическим показателям.
Кое-какие выводы.
1. Голые жеребята рождаются нормально развитыми, крупными, ярко выраженной породности.
ФГБНУ Всероссийский научно-исследовательский институт коневодства
п. Дивово, Рязанская обл., Россия
Заметные успехи в развитии молекулярно-генетических и информационных технологий дали мощный импульс для изучения геномов сельскохозяйственных животных, что позволило эффективно использовать на практике достижения маркер-вспомогательной и геномной селекции. Очевидный генетический прогресс в селекции животных значительной мере был обусловлен разработкой технологии полногеномного анализа, позволяющей выявлять локализацию генов, детерминирующих любой качественный или количественный признак, а затем изучать их строение и уровень полиморфизма на примере референтных популяций. Молекулярно-генетические методы типирования позволяют использовать для анализа любые ткани на любых стадиях онтогенеза и эффективно решать самые сложные вопросы генетической экспертизы. Типирование лошадей по локусам микросателлитов ДНК позволяет повысить эффективность генетический экспертизы происхождения практически до 100%, одновременно с этим изучать генетическую структуру и филогенетические связи пород, а также проводить мониторинг гетерогенности популяций (1, 2, 4, 7, 10).
Важной задачей является генетическая экспертиза племенной продукции на наличие дефектных генов, обуславливающих наследственные заболевания и снижение жизнеспособности. Сегодня у лошадей уже известны десятки генов, детерминирующих наследственные дефекты и заболевания, и разработаны молекулярно-генетические методы их диагностики. Выявлены ассоциации определенных генов с такими важными показателями работоспособности, как дистанционность и устойчивость рысистого аллюра.
Теоретические основы использования генетических маркеров (“сигналей”) в селекции животных были разработаны русским генетиком А.С. Серебровским еще в 30-х-40-годах прошлого века. Им были сформулированы основные требования, предъявляемые к генетическим маркерам - альтернативность, конкретная локализация на хромосомах и определенное влияние на изучаемый признак. Современные молекулярно-генетические технологии (включая полногеномное сканирование по чипам разной плотности) позволяют выполнить все эти условия. Более того, выявлены гены, достоверно влияющие на работоспособность и жизнеспособность лошадей, и у селекционеров уже имеется возможность применять геномную оценку лошадей (11, 12, 13, 14, 17).
Связь между аллельными генами и селекционируемыми признаками животных теоретически возможна через ряд генетических механизмов (3, 4). Ген, контролирующий образование белка или фермента, в силу своего плейотропного действия может одновременно влиять и на формирование полезного признака. Плейотропное действие гена может проявляться и как результат вторичного влияния выработанного под его контролем белка на отдельные биохимические и физиологические процессы в организме животного. Ввиду постоянства такой связи плейотропный эффект представляет большую ценность для практической селекции. Маркерные гены могут непосредственно детерминировать синтез белка (гормона, фермента), определяющего продуктивные качества, или находиться в одной группе сцепления с генами, определяющими хозяйственно-ценные признаки. В настоящее время исследователями активно ведется поиск генов-кандидатов, сцепленных с количественными признаками (QLT). Например, полиморфизм в локусе миостатина ( MSTN ) ассоциирован с дистанционностью лошадей чистокровной верховой породы (12, 15, 17), а ген DMRT 3 определяет склонность лошади двигаться иноходью или рысью (11).
При помощи модельного анализа было установлено (4), что отбор животных по генетическим маркерам эффективен даже при отсутствии плейотропии и сцепления. Последующие исследования подтвердили, что такие корреляции могут быть постоянными в ряде поколений, если они поддерживаются отбором, подбором и закрепляются инбридингом. В любом случае даже временные связи маркерных генов с хозяйственно- полезными признаками могут быть использованы в племенной работе с конкретными популяциями животных.
Благодаря быстрому внедрению ДНК-технологий и успешной работе по изучению генома лошади, общее число определяемых у лошадей маркерных генов уже превысило несколько сотен, что позволяет надежно контролировать значительную часть ее генома. Более того, систематическое тестирование всего поголовья лошадей основных заводских пород создает реальную основу для внедрения генетического мониторинга и других методов маркер-вспомогательной селекции в практику коневодства (5, 6, 8).
На основании обобщения результатов собственных исследований и анализа многочисленных публикаций по изучению и использованию полиморфизма структурных генов и участков ДНК ( STR , SNP ) лошадей, а также общей стратегии использования генетических маркеров в животноводстве, были сформулированы основные направления использования маркерной селекции в коневодстве (9).
1. Генетическая сертификация происхождения лошадей.
2. Изучение генетической структуры и филогенеза пород и популяций лошадей, включая оценку степени разнообразия и определение генетического сходства.
3.Проведение генетического мониторинга в породах лошадей, сохранение оригинальности и гетерогенности аллелофонда малочисленных пород.
4.Совершенствование метода линейного разведения, включая генетическую оценку степени дифференциации генеалогической структуры породы, определение генетического сходства с родоначальником.
5. Контроль применения родственного разведения, включая мониторинг
за нарастанием гомозиготности и оценку результатов инбридинга.
6. Селекция по маркерам, ассоциированным с хозяйственно-полезными
7. Диагностика наследственных дефектов и заболеваний, в том числе SCID, СА, HYPP и др.
7. Геномная селекция.
В отличие от ряда других отраслей животноводства, в которых широко практикуется крупномаштабная селекция, при племенной работе в коннозаводстве традиционно применяется индивидуальная система отбора и подбора, что является важной предпосылкой для внедрения методов маркерной селекции в повседневную коневодческую практику.
Генетические маркеры оказались незаменимым материалом для выявления диапазона популяционной и видовой изменчивости, изучения филогенеза, степени генетического сходства и последующей микроэволюции. пород лошадей. Благодаря многолетней работе лаборатории генетики ВНИИ коневодства по генетической сертификации лошадей проведена паспортизация всех отечественных конских пород по системам крови и микросателлитной ДНК. Среди всех изученных популяций своей генетической обособленностью как структурных генов, так и микросателлитной ДНК заметно выделялась орловская рысистая порода, представленная на дендрограмме (Рис. 1) центральной ветвью. Популяционный анализ подтвердил генетическую самобытность большинства конских пород и филогенетические связи между ними.
Изучение особенностей ядерной и митохондриальной ДНК лошадей разных пород и стран, а также найденных останков древних лошадей, позволили прояснить многие важные вопросы эволюции эквидов. В частности, была выявлена высокая вариабельность митохондриальной ДНК, свидетельствующая о наличии у домашних лошадей нескольких диких предков и существовании разных регионов одомашнения, одним из которых является территория Западной Сибири между Уральскими горами и рекой Обь.
Изучение генетических особенностей лошадей заводских и локальных пород лошадей стало основой для разработки методов генетического мониторинга в коневодстве. Мониторинг дает возможность дополнить традиционную селекцию новыми технологиями и позволяет вести отбор и подбор не только на фенотипическом, но и на генотипическом уровне. Одна из основных задач генетического мониторинга - это поддержание в породах генетического разнообразия, что является необходимым условием для творческой селекционной работы. Проведенный генетический мониторинг аллелофонда заводских пород лошадей показал, что за два десятилетия в большинстве пород наблюдалась тенденция снижения генетического разнообразия, что подтверждает динамика средних показателей уровня полиморфности и степени гетерозиготности полиморфных систем крови (6). Проведенный анализ показал, что низкий уровень генетической изменчивости ассоциируется с пониженными показателями плодовитости кобыл, поэтому потеря части аллелофонда в последующем может иметь непредсказуемые и негативные последствия для селекции.

Рис. 1. Дендрограмма генетических дистанций между породами лошадей по локусам микросателлитов ДНК.
Основным методом совершенствования пород лошадей является разведение по линиям, при этом традиционные зоотехнические методы оценки дифференциации сложившейся генеалогической структуры могут быть дополнены полученной генетической информацией. Изучение генетической структуры основных заводских пород лошадей по структурным генам показало наличие четко выраженных межлинейных различий по наличию и частоте встречаемости отдельных аллелей исследуемых локусов (7). Использование полиморфных локусов систем крови и микросателлитов ДНК при изучении генетических особенностей внутрипородных структур дает селекционерам дополнительную и весьма ценную информацию о степени генетического различия (или сходства) мужских и женских линий и позволяет наполнить родословную конкретной генетической информацией, которая может быть творчески использована при подборе и отборе.
Необходимо отметить, что разведение по линиям пока еще сводится к разведению по родословным. Как указывал Д.А. Кисловский (1965), большее или меньшее насыщение родословной кличками определенных животных не всегда соответствует их фактическому влиянию на пробанда, то есть обязательно необходимо принимать во внимание вероятностный характер коэффициентов, характеризующих степень генетического сходства на основании степени родства. Генетические маркеры являются именно тем критерием, который позволяет решать две важнейшие проблемы инбридинга: проводить оценку результатов родственного разведения по фактическому генотипическому сходству с инбридируемым предком и контролировать уровень гомозиготности у инбредных животных. При изучении влияния родственного разведения на гомозиготность орловских рысаков было установлено, что увеличение степени инбридинга не всегда сопровождается увеличением гомозиготности инбредных лошадей по полиморфным системам крови (8). Тенденция нарастания гомозиготности была отмечена только при близкородственном спаривании при коэффициенте инбридинга 5,0% и выше, фактически на границе проявления инбредной депрессии (Рис. 2).

Рис. 2. Степень гомозиготности лошадей орловской рысистой породы с разным уровнем инбридинга.
Несомненным достижением современной генетики является разработка методов ДНК-типирования десятков наследственных заболеваний лошадей, таких как тяжелый комбинированный иммунодефицит ( SCID ), мозжечковая атаксия (СА), несовершенный эпителиогенез ( JEB ), гиперкалимический паралич ( HYPP ), полисахаридная миопатия (PSSM), летальная белая масть (OLWS) и другие, как правило, вызванные различными дефектами нормальных генов (9, 16).
Таблица 1. Список однолокусных дефектов и болезней лошадей, для которых разработаны тест-системы
Выявить закономерности наследования особенностей экстерьера крайне сложно, так как на их развитие очень сильно влияют среда, условия выращивания молодняка. При неблагоприятных условиях у лошадей нарушается нормальная функция наследственных задатков, в результате формируются лошади большеголовые и грубоголовые, с короткими тонкими шеями, с недостаточно развитым корпусом (укороченные, с небольшим обхватом груди), со свислым крупом, узкой грудью, разметом ног, сближенностью в скакательных суставах, беднокостные, с плохо развитыми суставами, склонными к различного рода разращениям.

Наследование некоторых особенностей экстерьера
При нормальных условиях выращивания конского молодняка многими исследователями выявлены закономерности наследования некоторых особенностей телосложения у лошадей:
1. Сильно выраженные щетки доминируют над слабо выраженными. Развитие щеток определяется, по-видимому, несколькими факторами неполного доминирования.
2. При скрещивании лошадей шаговых пород с лошадьми быстроаллюрных пород обнаруживается неполное доминирование признаков тяжеловозов, а именно: помеси, как правило, имеют грубую голову, массивный корпус, часто раздвоенный круп, короткую мясистую шею; широкотелость, выраженная в индексе
| обхват груди | 100, |
| высота в холке |
доминирует над узкотелостью. Однако такой тип наследования можно наблюдать только в том случае, когда матери принадлежат к тяжеловозной, а отцы - к быстроаллюрной породе. При реципрокном скрещивании, то есть когда матери узкотелые, а отцы широкотелые, такой четкой закономерности не наблюдается. В этом случае обнаруживается преимущественное влияние материнского организма, усиленное влиянием фактора доминирования приспособленного типа.
Во всех случаях наблюдается доминирование длинноногости над коротконогостью.
3. Размеры животных (высота в холке, обхват груди, вес) относятся к категории количественных признаков. У лошадей они наследуются промежуточно. Размеры потомства в массе соответствуют средним размерам родителей. Отклонение в ту или другую сторону может быть незначительным за счет неполного доминирования широкотелого типа и длинноногости, а также доминирования приспособленного типа и влияния материнского организма.
Даже при скрещивании кобылы пони с жеребцом породы шайр потомство имеет приблизительно промежуточные размеры.
4. Во всех наставлениях по коневодству и коннозаводству считаются наследственными такие пороки экстерьера, как курба, жабка, шпат и рорер. Были предложены и схемы наследования этих пороков, как правило, по однофакториальному рецессивному типу. Однако позднейшие исследования показали, что наследование этих пороков осуществляется более сложным путем. Лошади с пороком шпат не родятся. Он появляется в возрасте 1 1 /2 лет и старше вследствие перенапряжения скакательных суставов и нарушения целостности суставных поверхностей. Слабость сухожильно-связочного аппарата, неблагоприятное механическое строение суставов и конечностей тазового пояса, непрочность костяка благоприятствуют развитию порока. Шпат чаще встречается у лошадей сырой (рыхлой) конституции, склонных к костным разращениям, с неправильным строением скакательного сустава. Аналогично наследуется и такой порок, как жабка.
Курба - порок сухожильно-связочного аппарата. Наследуется она в виде предрасположения; чаще наблюдается у лошадей недоразвитых, с иксобразной постановкой и саблистостью задних ног, при слабо развитых и сырых скакательных суставах.
Курба у лошади признак общей слабости сухожилий и связок.
Данных о наследовании рорера, или свистящего удушья, у лошадей нет, так как этот порок встречается не часто и обычно "хрипунов" не используют для воспроизводства.
У лошадей описаны наследственные дефекты, обусловленные летальными генами, которые вызывают гибель животного на разных стадиях роста и развития (до полового созревания). Вот их перечень: кривая шея, отсутствие передних ног или изуродованные (деформированные) передние ноги, полное отсутствие волос (кроющих и защитных), частичное отсутствие кожи, непроходимость прямой кишки, атаксия жеребят, выражающаяся в судорогах и параличах. Жеребята, отягощенные летальными задатками этих признаков, или родятся мертвыми, или гибнут в первые дни жизни. По общему мнению исследователей, каждый из летальных дефектов обусловлен одним рецессивным геном и проявляется только при гомозиготности по этому задатку.
Более часто чем летальные встречаются нелетальные наследственные дефекты у лошадей, такие как укороченная нижняя, а иногда верхняя челюсти (обусловлены одним рецессивным задатком), отсутствие радужной оболочки глаза, плоское копыто (обусловлены одним доминантным задатком), ярко выраженная коротконогость. Последний признак был описан в потомстве орловского рысистого жеребца Ветрогона 2. 10, 2, рожд. 1927 г. (Бор - Валькирия). В 1946 г. от него и русской рысистой кобылы Лесть, 1942 г. (Транзит - Лихая), в Александровском конном заводе родился коротконогий гнедой жеребчик Лапоть. При скрещивании Лаптя с кабардинской кобылой нормального сложения получен также коротконогий жеребчик Лапоть П.
Летальные и нелетальные дефекты организма, обусловленные одним рецессивным задатком, проявляются только в тех случаях, когда этот задаток находится в гомозиготном состоянии.
Миопатия накопления полисахаридов у лошадей ( EPSM , PSSM , EPSSM ) - это наследственное заболевание накопления гликогена у лошадей, которое вызывает рабдомиолиз при физической нагрузке . В настоящее время известно, что он поражает следующие породы: Американские четверть лошади, Американские пейнтбольные лошади, теплокровные, початковые, пони из долин, чистокровные, арабские, нью-форестские пони и большое количество тяжелых пород лошадей. Хотя этот синдром неизлечим, его можно контролировать с помощью соответствующей диеты и физических упражнений. В настоящее время существует 2 подтипа, известных как PSSM типа 1 и PSSM типа 2.
Содержание
- 1 Патофизиология нарушений накопления гликогена и субтипирование PSSM
- 1.1 Тип 1 PSSM
- 1.2 Дефицит фермента разветвления гликогена
- 1.3 Тип 2 PSSM
- 2 Презентация
- 3 Клинические признаки
- 3.1 Изменчивость фенотипа и модифицирующие гены
- 4 Диагностика
- 5 Менеджмент
- 5.1 Влияние на метаболизм
- 5.2 Диета
- 5.3 Упражнение
- 6 Ссылки
Патофизиология нарушений накопления гликогена и субтипирование PSSM
Гликоген представляет собой молекулярный полимер из глюкозы (а полисахарид ) , используемой для хранения энергии, и имеет важное значение для поддержания уровня глюкозы гомеостаза в крови, а также для обеспечения энергии скелетных мышц и сокращения сердечной мышцы. Молекулы глюкозы связаны в линейные цепи α-1,4- гликозидными связями . Кроме того, ответвления глюкозы образуются вне цепи α-1,6-гликозидными связями. 2 молекулы глюкозы соединены в α-1,4-гликозидные связи ферментом, известным как гликогенсинтаза . Эта связь может быть разорвана амилазой, когда организм хочет расщепить гликоген на глюкозу для получения энергии. Фермент разветвления гликогена отвечает за необходимые α-1,6-гликозидные связи, необходимые для начала разветвления этих линейных цепей.
Любое нарушение этой системы приводит к болезни накопления гликогена . В настоящее время существует 2 подкатегории болезней накопления гликогена у лошадей: миопатия накопления полисахаридов 1 типа, дефицит фермента ветвления гликогена и миопатия накопления полисахаридов 2 типа.
Тип 1 PSSM
PSSM типа 1 вызывается аутосомно-доминантной генетической мутацией, известной как GSY1. Эта мутация вызывает повышенную регуляцию гликогенсинтазы и высокие уровни гликогенсинтазы по сравнению с ферментом ветвления гликогена (GBE). Это измененное соотношение гликогенсинтазы и GBE приводит к образованию молекул гликогена с длинными цепями и небольшим количеством разветвлений, что делает эти молекулы в некоторой степени устойчивыми к амилазе . Мутация GSY1 связана с измененным метаболизмом глюкозы (но нормальным метаболизмом гликогена), а также с накоплением высоких уровней гликогена и аномального полисахарида в мышцах лошади. Кроме того, у некоторых лошадей обнаружена чувствительность к инсулину, что улучшает усвоение глюкозы мышечными клетками и способствует избыточному хранению гликогена, которое уже повышено вследствие мутации GSY1.
Дефицит фермента, ответвляющего гликоген
Низкий уровень фермента разветвления гликогена приводит к состоянию, известному как дефицит фермента разветвления гликогена. Это состояние вызвано мутацией гена GBE1, ответственного за выработку фермента ветвления гликогена. Впоследствии молекулы гликогена производятся с несколькими ответвлениями, что значительно снижает количество невосстанавливающих концов, резко замедляя скорость, с которой молекула может быть синтезирована или разрушена. Это вызывает низкий уровень мышечного гликогена, который очень устойчив к амилазе. Это заболевание обычно наблюдается у жеребят четвероногой лошади и приводит к летальному исходу.
Тип 2 PSSM
PSSM 2 типа - это категория заболеваний, которые приводят к аномальному отложению гликогена в скелетных мышцах лошади, что не связано с мутациями в GSY1 или GBE1.
Презентация

PSSM наиболее распространен у американских четвертных лошадей и связанных с ними пород ( пейнтбольные лошади , аппалуза , аппендикс-четвертинки ), рабочих пород лошадей (особенно бельгийских проектных и першеронов ) и теплокровных пород. Показано, что у бельгийских призывников частота ПССМ составляет 36%. Другие породы , которые были диагностированы с PSSM включают Аравитянин , Lipizzaner , Морган , Мустанг , перуанский Пасо , Rocky Mountain Horse , Стандардбред , Tennessee Walking Horse , Thoroughbred и National Show Horse . Было высказано предположение, что мутация GSY1 принесла некоторую пользу трудолюбивым животным с некачественным рационом и теперь наносит вред представителям тех «бережливых» пород, которые управляются с умеренным или низким уровнем работы и диетами с высоким содержанием неструктурных углеводов. .
PSSM типа 1 (гомозиготный или гетерозиготный по мутации GSY1) чаще встречается у четвероногих и родственных им пород, а также у рабочих пород, тогда как PSSM типа 2 (отрицательный по мутации GSY1) чаще встречается у других пород, включая теплокровных. Половой пристрастия к заболеванию нет.
Клинические признаки
Лошади с PSSM 1 типа обычно выглядят нормальными в состоянии покоя, но проявляют признаки рабдомиолиза при физической нагрузке ("связывания"), такие как укороченный шаг, скованность, крепкая мускулатура, потливость, боль или нежелание выполнять упражнения, когда их просят выполнить легкую работу. Хотя эпизоды рабдомиолиза при физической нагрузке являются одним из наиболее частых признаков, связанных с пораженными лошадьми (примерно у 37% пораженных животных), другие общие признаки включают аномалии походки, хромоту , мышечную слабость, которая может привести к неспособности подняться, колики, такие как боль и фасцикуляция, атрофия и / или ригидность мышц (чаще всего наблюдаются в полуперепончатых, полусухожильных и длинных мышцах).
Эти клинические признаки обычно впервые становятся очевидными, когда лошадь начинает тренироваться в молодом возрасте; однако у пораженных лошадей гистологические изменения, соответствующие повреждению мышц, в возрасте одного месяца, а также могут наблюдаться повышения креатинкиназы (CK), фермента, который повышается при повреждении мышц. Сопутствующее заболевание, такое как респираторная или желудочно-кишечная инфекция, может привести к повышению CK и потенциально опасному для жизни рабдомиолизу даже без упражнений. Лошади с PSSM часто имеют устойчиво повышенный уровень CK в состоянии покоя, что отличает заболевание от рецидивирующего рабдомиолиза при физической нагрузке, при котором лошади имеют нормальные концентрации CK между эпизодами.
Изменчивость фенотипа и модифицирующих генов
Некоторые пораженные животные могут оставаться субклиническими, у других могут быть легкие признаки, не препятствующие спортивным достижениям, в то время как у некоторых лошадей будут клинические признаки, препятствующие любым принудительным упражнениям. В редких случаях лошади умирают от острых эпизодов рабдомиолиза. Причина такой изменчивости фенотипа до конца не выяснена. Темперамент, пол и телосложение не влияют на выраженность клинических признаков. Однако факторы окружающей среды, такие как диета и упражнения, гетерозиготность или гомозиготность лошади по мутированному аллелю GSY1, а также наличие модифицирующих генов, играют роль. Кроме того, у некоторых пораженных лошадей может быть PSSM типа 2, который вызывает различные клеточные изменения и, следовательно, различные фенотипические эффекты.
Одним из таких модифицирующих генов является RYR1, который отвечает за регуляцию кальция в мышечных клетках. Мутация RYR1 вызывает злокачественную гипертермию - редкое, но потенциально смертельное заболевание, обычно связанное с анестезией. В то время как мутация RYR1 редко встречается у лошадей, в том числе у общей популяции четвертичных лошадей, она гораздо чаще встречается у четвероногих лошадей с мутацией GSY1. Лошади с обеими мутациями с большей вероятностью будут иметь тяжелый фенотип PSSM, включая более высокие уровни креатинкиназы (КК) в крови , более тяжелую непереносимость физических упражнений, более тяжелые эпизоды рабдомиолиза (более частые фасцикуляции мышц, более частые эпизоды, не связанные с упражнениями. , острая смерть) и плохой ответ на лечение PSSM.
Кроме того, дефекты в генах GSY1 и SCNA4, ответственные за гиперкалиемический периодический паралич (HYPP) у четвероногих лошадей и родственных пород, были обнаружены у 14% холтерных лошадей. Комбинация обоих этих генов может вызвать тяжелый рабдомиолиз, если лошадь станет лежачей из-за атаки HYPP.
Диагностика
Генетический тест доступен для PSSM 1 типа. Для этого теста требуется образец крови или волос, и он менее инвазивен, чем биопсия мышц. Однако он может быть менее полезен для пород, которые чаще страдают от PSSM 2 типа, например для легких пород лошадей. Часто биопсия мышц рекомендуется лошадям с клиническими признаками PSSM, но имеющими отрицательные результаты на мутацию GYS1.
Биопсия мышцы может быть взята из полумембранозных или полутендинозных (подколенных сухожилий) мышц. Биопсия окрашивается на гликоген, и интенсивность поглощения пятна в мышце, а также наличие каких-либо включений помогает определить диагноз PSSM. Этот тест - единственный метод диагностики PSSM 2 типа. Лошади с PSSM типа 1 обычно имеют в 1,5-2 раза больше нормального уровня гликогена в их скелетных мышцах. В то время как аномалии, указывающие на повреждение мышц, можно увидеть на гистологических срезах мышц уже в возрасте 1 месяца, для развития аномального накопления полисахаридов может потребоваться до 3 лет.
Управление
Влияние на обмен веществ
Лошади с PSSM имеют повышенный уровень мышечного гликогена в состоянии покоя. Во время упражнений уровень гликогена истощается быстрее, чем это наблюдается у здоровых лошадей, и снижается до уровня, который считается нормальным для покоящейся лошади без PSSM. Это показывает, что метаболизм гликогена у этих животных действительно нормальный. Однако лошади PSSM синтезируют мышечный гликоген в два раза быстрее, чем нормальная лошадь, после прекращения упражнений, что приводит к повышению мышечного гликогена. Точный механизм аномального метаболизма глюкозы еще не установлен, но он может иметь сходство с дефицитом фосфофруктокиназы у людей.
Связанные с четвертью лошади породы с PSSM показывают чувствительность к инсулину, что улучшает усвоение глюкозы клетками, и эти лошади очищают кровь от глюкозы после еды быстрее, чем здоровые лошади. Это обеспечивает легкий доступ к глюкозе для мышц, которые затем могут использовать субстрат для производства гликогена. Дефект GYS1, который активирует фермент гликогенсинтазы, позволяет мышцам использовать эту глюкозу для быстрого производства гликогена для хранения в мышцах. Удивительно, но повышенной чувствительности к инсулину у упряжных пород лошадей не наблюдается.
Манипуляции с питанием и физическими упражнениями могут быть использованы для противодействия этим метаболическим изменениям. Приблизительно у 50% лошадей, которые придерживаются диетических рекомендаций, и у 90% лошадей, которые придерживаются как диетических рекомендаций, так и рекомендаций по физическим упражнениям, эпизоды рабдомиолиза при физической нагрузке практически отсутствуют.
Диета
Для большинства лошадей диета оказывает значительное влияние на выраженность клинических признаков. Было показано, что лошади PSSM, получавшие диеты с высоким содержанием неструктурных углеводов (NSC), которые стимулируют секрецию инсулина, имеют повышенную тяжесть рабдомиолиза при физических нагрузках. Текущие рекомендации для лошадей с PSSM включают диету с низким содержанием крахмала и высоким содержанием жиров. Диеты с низким содержанием крахмала вызывают низкий уровень глюкозы в крови и инсулина после еды, что может снизить количество глюкозы, поглощаемой мышечными клетками. Диеты с высоким содержанием жиров увеличивают концентрацию свободных жирных кислот в крови, что может способствовать использованию жира для получения энергии (посредством окисления свободных жирных кислот) по сравнению с метаболизмом глюкозы. Лошади с наиболее серьезными клиническими признаками часто демонстрируют наибольшее улучшение в диете.
Диетические рекомендации обычно включают сочетание ограничения калорий, снижения суточного содержания NSC и увеличения количества пищевых жиров. Рекомендации по диете должны быть сбалансированы с оценкой состояния тела животного и уровнем физических нагрузок, так как может быть полезно подождать увеличения количества диетического жира после того, как животное с ожирением похудеет. В рационе должно быть Упражнение
Лошади с PSSM проявляют меньше клинических признаков, если их физическая нагрузка постепенно увеличивается с течением времени (т.е. они медленно кондиционируются). Кроме того, у них гораздо больше шансов развить мышечную жесткость и рабдомиолиз, если они выполняются после длительного отдыха.
Обычно у лошадей меньше клинических признаков, когда их просят выполнять короткие периоды работы на максимальном уровне активности (анаэробные упражнения), хотя они испытывают трудности с достижением максимальной скорости и устают быстрее, чем здоровые лошади. У них больше повреждения мышц, когда их просят выполнять упражнения с меньшей интенсивностью в течение более длительного периода времени (аэробная активность) из-за дефицита энергии в мышцах.
Читайте также: