
Какие половые хромосомы у курицы
Опубликовано: 29.04.2024

Поговорим о том, сколько хромосом у петуха и курицы. Как и у млекопитающих, клетки этих птиц имеют женский или мужской хромосомный набор. Ученые долгое время утверждали, что невозможно определить пол эмбриона курочки до определенного момента развития. Но с помощью лабораторных исследований удалось выяснить, что это не так. Клетки зародыша хранят информацию о половой принадлежности уже с третьей недели формирования.
- О курицах и петухах
- Поговорим о хромосомах
- Где находятся
- За что отвечают
- Что определяет половую принадлежность птиц
О курицах и петухах
Куры – одни из распространенных жителей фермерских хозяйств. В благоприятных условиях они могут жить 12-15 лет. Но на практике такое встречается редко. Птицу забивают после 2-3 лет жизни, когда у нее падает яичная производительность. На крупных птицефабриках куриц отправляют на убой через год после первой кладки.
Средний вес самки – 3,5 кг, а яйценоскость — 120 яиц в год. Но производительность зависит от породы и условий содержания. Узнайте больше в статье «Что за птица домашняя курица».
Петух является хозяином курятника, знаменит своенравным характером и мужеством. Он главный зачинщики драк в стаде. Поэтому в курином семействе должен жить только один петушок. Иначе постоянно будут конфликты.

На каждого самца приходится около 10 курочек. Если их будет больше, начнутся проблемы со здоровьем и производительностью.
Главные отличия самцов от самок:
- длинный хвост;
- большие сережки;
- роскошное яркое оперение.
Гордостью петушков является мясистый алый гребень. Подробности в статье «Как должен выглядеть петух: описание птицы».

Поговорим о хромосомах
Где находятся
Это нуклеопротеидные структуры, располагающиеся в клетках организма птиц. Они являются носителями генетической информации, состоят из спиралеобразных молекул ДНК и белков.
Полный хромосомный набор кур называется кариотип. Он включает информацию о форме, размерах и численности генетического материала.
Хромосомы есть у всех живых организмов. Но у каждой птицы набор свой. Он постоянный и не меняется с возрастом.
Внешне структуры похожи на длинную нитку. На ней располагается множество бусинок – генов. Каждый ген занимает определенное место – локус.

За что отвечают
Гены никогда не передвигаются по хромосоме. Их задача – управление признаками индивидуума.
Хромосомы занимаются хранением и передачей накопленной информации от матери к потомству.
После многочисленных исследований ученые установили, сколько хромосом у курицы и петуха – 78. Это достаточно большое количество по сравнению с другими млекопитающими. Например, у людей их всего 46.
В ходе эволюции куры и петушки потерпели меньше всего генетических изменений, по сравнению с другими птицами.

Что определяет половую принадлежность птиц
78 хромосом бывает только у здоровых петушков или кур. Если во время формирования яиц у самки начнутся проблемы с развитием эмбриона, их количество может поменяться.
У кур XY – набор хромосом, у петухов – XX. У многих млекопитающих, в том числе человека, наоборот.
Ученые из Великобритании провели исследование здоровых эмбрионов кур. Выяснилось, что менее чем через день после оплодотворения определяется половая принадлежность эмбриона.
У других млекопитающих это происходит только после формирования репродуктивных желез. Данный факт удалось определить по выработке РНК-молекул.
Подробные инструкции об определении пола цыпленка до вылупления указаны в статье «Как можно определить пол цыпленка по яйцу».
Как узнать пол после вылупления читайте в статье «Петух или курица: как определять пол цыпленка».

Уважаемые фермеры! Если вы нашли полезную для себя информацию, поставьте, пожалуйста, лайк.
В комментариях поделитесь, пожалуйста, историями о том, как вы определяли пол цыплят.

CC BY
Аннотация научной статьи по биологическим наукам, автор научной работы — Тагиров М. Т.
Рассмотрены современные представления о генетических основах детерминации пола у птиц . Подчиняясь генотипическому механизму определения пола, птицы реализуют сложную систему взаимодействий половых хромосом. Z -хромосома содержит дозозависимую детерминанту мужского пола — ген DMTR1, обеспечивающий развитие семенников при наличии двух копий в комбинации ZZ. Еще не идентифицированные факторы на W -хромосоме, с одной стороны, подавляют активность гена DMTR1, а с другой — активируют энзим ароматазу, что в комбинации ZW способствует развитию женского пола. Выдвинуто предположение, согласно которому по мере появления в ряду различных классов животных приспособлений, обеспечивающих стабильность зародышевого развития, снижается зависимость половой детерминации от внешних условий и внутренних сигналов на поздних стадиях эмбриогенеза. В то же время появляются закрепленные на генетическом уровне половые детерминанты, активные на самых ранних стадиях развития.
Похожие темы научных работ по биологическим наукам , автор научной работы — Тагиров М. Т.
SEX DETERMINATION AND CONTROL MECHANISMS IN BIRDS
The modern conception of genetic bases of bird ’s sex determination is considered. Following the genotypic mechanism of sex determination , birds implement a complex system of sex chromosome relationships. Z chromosome contains a dose-dependent male sex determinant — gene DMTR1 ensuring the testicles development upon two copy availability in the ZZ combination. Some factors on the W chromosome are not yet identified. On the one hand, they suppress the activity of the DMTR1 gene and on the other hand they activate aromatase enzyme, which in ZW combination promotes the female sex deve lopment. It was suggested that while a number of different classes of animal adaptations are appeared ensuring stable embryo development, the dependence of sex determination upon environment and inner signals at the later stages of embryogenesis is reduced. At the same time, sex determinants fixed on genetic level are appeared that are active at the very early stages of deve lopment.
Текст научной работы на тему «Механизмы контроля и детерминации пола у птиц»
МЕХАНИЗМЫ КОНТРОЛЯ И ДЕТЕРМИНАЦИИ ПОЛА У ПТИЦ
1Институт птицеводства НААН Украины, с. Борки, Харьковская обл.
Рассмотрены современные представления о генетических основах детерминации пола у птиц. Подчиняясь генотипическому механизму определения пола, птицы реализуют сложную систему взаимодействий половых хромосом. Z-хромосома содержит дозозависимую детерминанту мужского пола — ген DMTR1, обеспечивающий развитие семенников при наличии двух копий в комбинации ZZ. Еще не идентифицированные факторы на W-хромосоме, с одной стороны, подавляют активность гена DMTR1, а с другой — активируют энзим ароматазу, что в комбинации ZW способствует развитию женского пола. Выдвинуто предположение, согласно которому по мере появления в ряду различных классов животных приспособлений, обеспечивающих стабильность зародышевого развития, снижается зависимость половой детерминации от внешних условий и внутренних сигналов на поздних стадиях эмбриогенеза. В то же время появляются закрепленные на генетическом уровне половые детерминанты, активные на самых ранних стадиях развития.
Ключевые слова: птица, детерминация пола, половые хромосомы, инверсия пола, мейотический драйв, гонады.
Проявляемый в последнее время повышенный интерес к половой детерминации у птиц объясняется прежде всего тем, что решение проблемы ранней диагностики пола или управления половой детерминацией птиц позволит сэкономить значительные финансовые, энергетические и пищевые ресурсы. Появится возможность инкубации либо производства зародышей определенного пола [1, 2]. В странах с высоким уровнем жизни все чаще затрагивается этическая сторона проблемы, связанная с массовым умерщвлением цыплят (в частности, петушков) в яичном птицеводстве, что связано с неэкономичностью их выращивания на мясо [1, 3]. В связи с этим вполне закономерны успехи, достигнутые за последние 20-30 лет в понимании молекулярно-генетических основ регуляции половой дифференциации у позвоночных животных 4.
Целью настоящей статьи является краткий обзор современных представлений о генетических механизмах определения и регуляции половой детерминации у птиц.
Генетические механизмы определения пола у птиц
В отличие от млекопитающих, у птиц гетерогаметным полом является самка
с хромосомной конституцией ZW. У самцов половые хромосомы представлены гомога-метной парой ZZ. Окончательный механизм детерминации пола у птиц до сих пор не выяснен, хотя в последнее время получены убедительные экспериментальные данные, в значительной степени проясняющие эту ситуацию 6.
Первый, наиболее простой механизм детерминации пола, аналогичный тому, что действует у млекопитающих, — это предположение о наличии на W-хромосоме фактора, детерминирующего самку. W-хромосома похожа на У-хромосому млекопитающих: маленького размера, содержит мало активных генов и много повторяющейся ДНК. Считают, что половые хромосомы птиц произошли от аутосомной пары, а W — это деградированная Z-хромосома [7].
Неравномерное распределение хромосом у разных полов обусловливает разность в дозе генов, которые могут привести к дефектам развития. Зарегистрированы различные отклонения у человека, вызванные анеуплоидией по разным хромосомам: синдром Дауна — трисомия по 21-й хромосоме, синдром Эдварда — трисомия по 18-й хромосоме, синдром Кляйнфельтера — 47-й, ХХУ и синдром Тернера — 45-й, Х. Поскольку на ранних стадиях развития
зародыш млекопитающих весьма чувствителен к хромосомным отклонениям, только небольшая часть эмбрионов с хромосомными отклонениями доживают до рождения [8]. Известно, что независимо от количества Х-хромосом присутствие Y-хромосомы у млекопитающих индуцирует тип развития самца: XXY — особи являются самцами, а ХО — самками. К сожалению, у птиц не зарегистрированы анеуплоидные формы по половым хромосомам и их считают генетическими леталями [9].
В то же время птицы менее чувствительны к отклонениям количества хромосом; зарегистрировано множество фактов живых триплоидов или триплоид-диплоид мозаиков [10, 11]. Так, в коммерческих стадах 1 из 2 000 несушек является интерсексом, имеющим промежуточные свойства самца и самки. Большинство из этих интерсексов — либо триплоиды ААА/ZZW, либо триплоид-диплоид мозаики. Описаны живые особи триплоидов ZZZ и ZZW, но, к сожалению, они всегда не фертильны. Мозаики (AAA/ZZZ, AA/ZZ) характеризуются отставанием в эмбриональном развитии, пониженной митотической активностью [12]. Анализ триплоидов (ZZW) показал, что они являются интер-сексами [13]. Будучи по фенотипу самками при рождении, триплоиды имеют правый семенник, левый овотестис, а к возрасту полового созревания женская составляющая овотестиса дегенерирует и птицы становятся самцами. Можно предположить присутствие на W-хромосоме детерминанты самки, которая находится под дозозависимым влиянием Z-хромосомы, и эта детерминанта не носит доминантный характер [14].
Анализ химер птиц типа ZZ ZW по -казал, что первичные половые клетки ZZ способны пройти мейоз в тканях ZW яичника и формировать функциональные ооциты в отсутствие W-хромосомы [15]. Таким образом, можно сделать вывод, что присутствие W-хромосомы не является обязательным условием для прохождения мейоза, и что гены оогенеза, вероятнее всего, расположены на Z-хромосоме или аутосомах.
Разные виды животных выработали различные реакции на дозу генов хромосом. Млекопитающие контролируют дозу генов половой хромосомы через инактивацию одной Х-хромосомы в каждой клетке самки в комбинации с усилением экспрессии генов в оставшейся Х-хромосоме обеих полов [16].
Вопрос, касающийся компенсации дозы генов и половой детерминации у птиц усложняется тем, что у бескилевых птиц практически нет кариотипических различий по Z- и W-хромосомам [17]. А такие таксономоческие группы птиц, как соколообразные (Falconiformes), журавлеобразные (Gruiformes) и дятлеобразные (Piciformes) имеют сильно увеличенные половые хромосомы (как Z, так и W), делая Z самой крупной хромосомой кариотипа [18]. Связано ли это с увеличением количества генов либо их плотности, еще не установлено.
На эухроматиновом плече W-хромосомы птиц были открыты два гена, которые могли бы выступить в роли половых детерминант. Первая из них — ген ASW [19]. Этот ген экспрессируется только в эмбрионах женского пола, в основном в гонадах, в то время как его единственный гомолог на Z-хромосоме — ZPKCI — экспрессируется в значительно меньшей степени у обоих полов [19]. Однако, несмотря на многократные повторы гена ASW на W-хромосоме, половое развитие триплоидов ZZW показывает, что активность этого гена недостаточна для индукции перманентного развития таких особей по пути формирования самки. Предположение об определяющей роли гена ASW в детерминации яичника птиц опровергается и тем обстоятельством, что активность этого гена зафиксирована и в некоторых других органах, кроме гонад [20]. ASW также оказался неспособным индуцировать развитие самки при чрезмерной его экспрессии в эмбрионах самцов [4]. Кроме того, этот ген не обнаружен у бескилевых птиц [19]. У них половые хромосомы гомоморфны, но так же, как и у других представителей класса им присуща генотипическая детерминация пола и, вероятнее всего, общий с другими птицами генетический триггер.
Второй ген — FET1 — расположен на эухроматиновом участке короткого плеча W-хромосомы, не связан с ASW-геном и не имеет гомолога на Z-хромосоме. Он экспрессируется исключительно в мочеполовой системе самки в период, предшествующий половой дифференциации (4,5-6,5 сут; 25-30-я стадии развития). Уровень экспрессии FET1 в левой гонаде значительно выше, чем в правой. Этот ген не проявляет гомологии с каким-либо из известных генов и кодирует протеин из 434 аминокислотных остатков с предполагаемой сигнальной последовательностью и трансмембранным доменом. У млекопитающих нет ортологов генов ASW и FET1 [21].
Даже если эти гены окажутся детерминантами яичников у птиц, еще остаются отсутствующие звенья на пути активации ароматазы, ключевого энзима, превращающего тестостерон в эстрадиол. ASW/WPKCI и FETl экспрессируются раньше, чем проявляется активность ароматазы. Именно в связи с этим активация этого гена невозможна без участия других факторов [21].
Вторая гипотеза предполагает детерминацию мужского пола птиц двойной дозой генов, сцепленных с Z-хромосомой. Необходимым условием для этого является то, что такой ген не должен подвергаться компенсаторному механизму, согласно которому гены на одной из Z-хромосом в комбинации ZZ инактивируются. Хотя активность многих генов на Z-хромосоме у самок и самцов подобна [22], есть данные, что некоторые мРНК транскрибируются с генов обеих Z-хромосом в клетках самцов [23]. Поэтому, если у птиц и происходит компенсация дозы генов в ходе раннего развития, то этому механизму подчинены не все гены на Z-хромосоме.
Одним из генов, не подверженных компенсации дозы, является ген DMRTl — наиболее подходящий кандидат на роль детерминанты мужского пола у птиц. Он находится на Z-хромосоме, но не имеет гомолога на W-хромосоме. DMRTl экспрессируется в гонадах птиц до и в ходе их дифференциации, будучи более активным в семенниках, чем в яичниках [24, 25]. Протеин DMRTl изначально находится в ядрах клеток медуллярных канальцев, где начинается управление процессом формирования семенников. Более того, в клетках генетических самок, у которых произошла реверсия пола под влиянием ингибитора ароматазы, активность DMRTl возрастает, несмотря на присутствие одной копии [26]. DMRTl — пример консервативности полоопределяющих генов у различных типов животных, поскольку его аналоги задействованы в половой дифференциации как млекопитающих, так и насекомых [27].
Согласно данным Smith et al., подтверждающим дозозависимую гипотезу половой детерминации у птиц, высокие дозы гена DMRTl инициируют дифференциацию семенников у самцов, активируя экспрессию гена SOX9 и подавляя активность ароматазы [4]. Ген DMRTl полностью отвечает требованиям к половой детерминанте птиц. Этот ген, сцепленный с полом, находится на Z-хромосоме у всех птиц, включая бескилевых [28]. Он экспрессируется исключительно в мочеполовой
системе перед половой дифференциацией гонад эмбрионов птиц. У других позвоноч -ных DMRT1 также участвует в развитии семенников. Было показано, что у нокаут -ных по гену DMRT1 мышей наблюдается нарушение постнатального развития семенников, а при микроделециях последовательности этого гена у человека наблюдается тестикулярный дисгенез. У рептилий с температурной детерминацией пола концентрация DMRT1 увеличивается в термочувствительный период и только при температуре, при которой происходит определение пола самца. [29, 30]. У рыбы медака Oryzias latipes двойная копия DMRT1, dmy/dmrtlb, является ведущей детерминантой семенников, а у амфибий копия DMRT1, сцепленная с W, dmw определяет развитие яичников
[31]. Таким образом, DMRT1 может оказаться дозочувствительным наследственным половым детерминантом как у птиц, так и у низших позвоночных, который заменяется геном SRY у млекопитающих.
Новая модель половой детерминации эмбрионов кур включает непосредственное взаимодействие фактора, производного W-хромосомы, и Z-хромосомы. Teranishi et al.
[32] обнаружили на коротком плече Z-хромосомы гиперметилированный участок (МНМ-участок), состоящий из более чем 200 копий тандемных повторов размером 2,2 kb. Этот участок гиперметилирован и транскрипцион-но неактивен на обеих Z-хромосомах самцов, в то время как у самок он деметилирован, и продукт его транскрипции — высокомолекулярная РНК — ингибирует прилегающий DMRT1 -локус. Интересно, что в триплоидах ZZZ все три Z-хромосомы гиперметилирова-ны и неактивны, в то время как в триплоидах ZZW МНМ-участок гипометилирован и транскрибируется на обеих Z-хромосомах [32]. Эти данные могут свидетельствать о том, что гены на W-хромосоме могут быть вовлечены в процесс деметилирования МНМ-участка Z-хромосомы. У ZZ-самцов в отсутствие W-хромосомы МНМ-участок остается гипер-метилированным и неактивным, не синтезируется РНК и не происходит подавление активности DMRT1 (рис. 1).
Если предположить, что роль фактора F выполняет продукт гена FET1, то результатом подавления его активности должен быть эффект, наблюдаемый при ингибировании активности ароматазы, т. е. реверсия пола в сторону самца. Нокдаун гена FET1 позволил бы прояснить это предположение, как это было показано с геном DMRT1 [4].
CC BY
Аннотация научной статьи по биологическим наукам, автор научной работы — Тагиров М. Т.
Рассмотрены современные представления о генетических основах детерминации пола у птиц . Подчиняясь генотипическому механизму определения пола, птицы реализуют сложную систему взаимодействий половых хромосом. Z -хромосома содержит дозозависимую детерминанту мужского пола — ген DMTR1, обеспечивающий развитие семенников при наличии двух копий в комбинации ZZ. Еще не идентифицированные факторы на W -хромосоме, с одной стороны, подавляют активность гена DMTR1, а с другой — активируют энзим ароматазу, что в комбинации ZW способствует развитию женского пола. Выдвинуто предположение, согласно которому по мере появления в ряду различных классов животных приспособлений, обеспечивающих стабильность зародышевого развития, снижается зависимость половой детерминации от внешних условий и внутренних сигналов на поздних стадиях эмбриогенеза. В то же время появляются закрепленные на генетическом уровне половые детерминанты, активные на самых ранних стадиях развития.
Похожие темы научных работ по биологическим наукам , автор научной работы — Тагиров М. Т.
SEX DETERMINATION AND CONTROL MECHANISMS IN BIRDS
The modern conception of genetic bases of bird ’s sex determination is considered. Following the genotypic mechanism of sex determination , birds implement a complex system of sex chromosome relationships. Z chromosome contains a dose-dependent male sex determinant — gene DMTR1 ensuring the testicles development upon two copy availability in the ZZ combination. Some factors on the W chromosome are not yet identified. On the one hand, they suppress the activity of the DMTR1 gene and on the other hand they activate aromatase enzyme, which in ZW combination promotes the female sex deve lopment. It was suggested that while a number of different classes of animal adaptations are appeared ensuring stable embryo development, the dependence of sex determination upon environment and inner signals at the later stages of embryogenesis is reduced. At the same time, sex determinants fixed on genetic level are appeared that are active at the very early stages of deve lopment.
Текст научной работы на тему «Механизмы контроля и детерминации пола у птиц»
МЕХАНИЗМЫ КОНТРОЛЯ И ДЕТЕРМИНАЦИИ ПОЛА У ПТИЦ
1Институт птицеводства НААН Украины, с. Борки, Харьковская обл.
Рассмотрены современные представления о генетических основах детерминации пола у птиц. Подчиняясь генотипическому механизму определения пола, птицы реализуют сложную систему взаимодействий половых хромосом. Z-хромосома содержит дозозависимую детерминанту мужского пола — ген DMTR1, обеспечивающий развитие семенников при наличии двух копий в комбинации ZZ. Еще не идентифицированные факторы на W-хромосоме, с одной стороны, подавляют активность гена DMTR1, а с другой — активируют энзим ароматазу, что в комбинации ZW способствует развитию женского пола. Выдвинуто предположение, согласно которому по мере появления в ряду различных классов животных приспособлений, обеспечивающих стабильность зародышевого развития, снижается зависимость половой детерминации от внешних условий и внутренних сигналов на поздних стадиях эмбриогенеза. В то же время появляются закрепленные на генетическом уровне половые детерминанты, активные на самых ранних стадиях развития.
Ключевые слова: птица, детерминация пола, половые хромосомы, инверсия пола, мейотический драйв, гонады.
Проявляемый в последнее время повышенный интерес к половой детерминации у птиц объясняется прежде всего тем, что решение проблемы ранней диагностики пола или управления половой детерминацией птиц позволит сэкономить значительные финансовые, энергетические и пищевые ресурсы. Появится возможность инкубации либо производства зародышей определенного пола [1, 2]. В странах с высоким уровнем жизни все чаще затрагивается этическая сторона проблемы, связанная с массовым умерщвлением цыплят (в частности, петушков) в яичном птицеводстве, что связано с неэкономичностью их выращивания на мясо [1, 3]. В связи с этим вполне закономерны успехи, достигнутые за последние 20-30 лет в понимании молекулярно-генетических основ регуляции половой дифференциации у позвоночных животных 4.
Целью настоящей статьи является краткий обзор современных представлений о генетических механизмах определения и регуляции половой детерминации у птиц.
Генетические механизмы определения пола у птиц
В отличие от млекопитающих, у птиц гетерогаметным полом является самка
с хромосомной конституцией ZW. У самцов половые хромосомы представлены гомога-метной парой ZZ. Окончательный механизм детерминации пола у птиц до сих пор не выяснен, хотя в последнее время получены убедительные экспериментальные данные, в значительной степени проясняющие эту ситуацию 4.
Первый, наиболее простой механизм детерминации пола, аналогичный тому, что действует у млекопитающих, — это предположение о наличии на W-хромосоме фактора, детерминирующего самку. W-хромосома похожа на У-хромосому млекопитающих: маленького размера, содержит мало активных генов и много повторяющейся ДНК. Считают, что половые хромосомы птиц произошли от аутосомной пары, а W — это деградированная Z-хромосома [7].
Неравномерное распределение хромосом у разных полов обусловливает разность в дозе генов, которые могут привести к дефектам развития. Зарегистрированы различные отклонения у человека, вызванные анеуплоидией по разным хромосомам: синдром Дауна — трисомия по 21-й хромосоме, синдром Эдварда — трисомия по 18-й хромосоме, синдром Кляйнфельтера — 47-й, ХХУ и синдром Тернера — 45-й, Х. Поскольку на ранних стадиях развития
зародыш млекопитающих весьма чувствителен к хромосомным отклонениям, только небольшая часть эмбрионов с хромосомными отклонениями доживают до рождения [8]. Известно, что независимо от количества Х-хромосом присутствие Y-хромосомы у млекопитающих индуцирует тип развития самца: XXY — особи являются самцами, а ХО — самками. К сожалению, у птиц не зарегистрированы анеуплоидные формы по половым хромосомам и их считают генетическими леталями [9].
В то же время птицы менее чувствительны к отклонениям количества хромосом; зарегистрировано множество фактов живых триплоидов или триплоид-диплоид мозаиков [10, 11]. Так, в коммерческих стадах 1 из 2 000 несушек является интерсексом, имеющим промежуточные свойства самца и самки. Большинство из этих интерсексов — либо триплоиды ААА/ZZW, либо триплоид-диплоид мозаики. Описаны живые особи триплоидов ZZZ и ZZW, но, к сожалению, они всегда не фертильны. Мозаики (AAA/ZZZ, AA/ZZ) характеризуются отставанием в эмбриональном развитии, пониженной митотической активностью [12]. Анализ триплоидов (ZZW) показал, что они являются интер-сексами [13]. Будучи по фенотипу самками при рождении, триплоиды имеют правый семенник, левый овотестис, а к возрасту полового созревания женская составляющая овотестиса дегенерирует и птицы становятся самцами. Можно предположить присутствие на W-хромосоме детерминанты самки, которая находится под дозозависимым влиянием Z-хромосомы, и эта детерминанта не носит доминантный характер [14].
Анализ химер птиц типа ZZ ZW по -казал, что первичные половые клетки ZZ способны пройти мейоз в тканях ZW яичника и формировать функциональные ооциты в отсутствие W-хромосомы [15]. Таким образом, можно сделать вывод, что присутствие W-хромосомы не является обязательным условием для прохождения мейоза, и что гены оогенеза, вероятнее всего, расположены на Z-хромосоме или аутосомах.
Разные виды животных выработали различные реакции на дозу генов хромосом. Млекопитающие контролируют дозу генов половой хромосомы через инактивацию одной Х-хромосомы в каждой клетке самки в комбинации с усилением экспрессии генов в оставшейся Х-хромосоме обеих полов [16].
Вопрос, касающийся компенсации дозы генов и половой детерминации у птиц усложняется тем, что у бескилевых птиц практически нет кариотипических различий по Z- и W-хромосомам [17]. А такие таксономоческие группы птиц, как соколообразные (Falconiformes), журавлеобразные (Gruiformes) и дятлеобразные (Piciformes) имеют сильно увеличенные половые хромосомы (как Z, так и W), делая Z самой крупной хромосомой кариотипа [18]. Связано ли это с увеличением количества генов либо их плотности, еще не установлено.
На эухроматиновом плече W-хромосомы птиц были открыты два гена, которые могли бы выступить в роли половых детерминант. Первая из них — ген ASW [19]. Этот ген экспрессируется только в эмбрионах женского пола, в основном в гонадах, в то время как его единственный гомолог на Z-хромосоме — ZPKCI — экспрессируется в значительно меньшей степени у обоих полов [19]. Однако, несмотря на многократные повторы гена ASW на W-хромосоме, половое развитие триплоидов ZZW показывает, что активность этого гена недостаточна для индукции перманентного развития таких особей по пути формирования самки. Предположение об определяющей роли гена ASW в детерминации яичника птиц опровергается и тем обстоятельством, что активность этого гена зафиксирована и в некоторых других органах, кроме гонад [20]. ASW также оказался неспособным индуцировать развитие самки при чрезмерной его экспрессии в эмбрионах самцов [4]. Кроме того, этот ген не обнаружен у бескилевых птиц [19]. У них половые хромосомы гомоморфны, но так же, как и у других представителей класса им присуща генотипическая детерминация пола и, вероятнее всего, общий с другими птицами генетический триггер.
Второй ген — FET1 — расположен на эухроматиновом участке короткого плеча W-хромосомы, не связан с ASW-геном и не имеет гомолога на Z-хромосоме. Он экспрессируется исключительно в мочеполовой системе самки в период, предшествующий половой дифференциации (4,5-6,5 сут; 25-30-я стадии развития). Уровень экспрессии FET1 в левой гонаде значительно выше, чем в правой. Этот ген не проявляет гомологии с каким-либо из известных генов и кодирует протеин из 434 аминокислотных остатков с предполагаемой сигнальной последовательностью и трансмембранным доменом. У млекопитающих нет ортологов генов ASW и FET1 [21].
Даже если эти гены окажутся детерминантами яичников у птиц, еще остаются отсутствующие звенья на пути активации ароматазы, ключевого энзима, превращающего тестостерон в эстрадиол. ASW/WPKCI и FETl экспрессируются раньше, чем проявляется активность ароматазы. Именно в связи с этим активация этого гена невозможна без участия других факторов [21].
Вторая гипотеза предполагает детерминацию мужского пола птиц двойной дозой генов, сцепленных с Z-хромосомой. Необходимым условием для этого является то, что такой ген не должен подвергаться компенсаторному механизму, согласно которому гены на одной из Z-хромосом в комбинации ZZ инактивируются. Хотя активность многих генов на Z-хромосоме у самок и самцов подобна [22], есть данные, что некоторые мРНК транскрибируются с генов обеих Z-хромосом в клетках самцов [23]. Поэтому, если у птиц и происходит компенсация дозы генов в ходе раннего развития, то этому механизму подчинены не все гены на Z-хромосоме.
Одним из генов, не подверженных компенсации дозы, является ген DMRTl — наиболее подходящий кандидат на роль детерминанты мужского пола у птиц. Он находится на Z-хромосоме, но не имеет гомолога на W-хромосоме. DMRTl экспрессируется в гонадах птиц до и в ходе их дифференциации, будучи более активным в семенниках, чем в яичниках [24, 25]. Протеин DMRTl изначально находится в ядрах клеток медуллярных канальцев, где начинается управление процессом формирования семенников. Более того, в клетках генетических самок, у которых произошла реверсия пола под влиянием ингибитора ароматазы, активность DMRTl возрастает, несмотря на присутствие одной копии [26]. DMRTl — пример консервативности полоопределяющих генов у различных типов животных, поскольку его аналоги задействованы в половой дифференциации как млекопитающих, так и насекомых [27].
Согласно данным Smith et al., подтверждающим дозозависимую гипотезу половой детерминации у птиц, высокие дозы гена DMRTl инициируют дифференциацию семенников у самцов, активируя экспрессию гена SOX9 и подавляя активность ароматазы [4]. Ген DMRTl полностью отвечает требованиям к половой детерминанте птиц. Этот ген, сцепленный с полом, находится на Z-хромосоме у всех птиц, включая бескилевых [28]. Он экспрессируется исключительно в мочеполовой
системе перед половой дифференциацией гонад эмбрионов птиц. У других позвоноч -ных DMRT1 также участвует в развитии семенников. Было показано, что у нокаут -ных по гену DMRT1 мышей наблюдается нарушение постнатального развития семенников, а при микроделециях последовательности этого гена у человека наблюдается тестикулярный дисгенез. У рептилий с температурной детерминацией пола концентрация DMRT1 увеличивается в термочувствительный период и только при температуре, при которой происходит определение пола самца. [29, 30]. У рыбы медака Oryzias latipes двойная копия DMRT1, dmy/dmrtlb, является ведущей детерминантой семенников, а у амфибий копия DMRT1, сцепленная с W, dmw определяет развитие яичников
[31]. Таким образом, DMRT1 может оказаться дозочувствительным наследственным половым детерминантом как у птиц, так и у низших позвоночных, который заменяется геном SRY у млекопитающих.
Новая модель половой детерминации эмбрионов кур включает непосредственное взаимодействие фактора, производного W-хромосомы, и Z-хромосомы. Teranishi et al.
[32] обнаружили на коротком плече Z-хромосомы гиперметилированный участок (МНМ-участок), состоящий из более чем 200 копий тандемных повторов размером 2,2 kb. Этот участок гиперметилирован и транскрипцион-но неактивен на обеих Z-хромосомах самцов, в то время как у самок он деметилирован, и продукт его транскрипции — высокомолекулярная РНК — ингибирует прилегающий DMRT1 -локус. Интересно, что в триплоидах ZZZ все три Z-хромосомы гиперметилирова-ны и неактивны, в то время как в триплоидах ZZW МНМ-участок гипометилирован и транскрибируется на обеих Z-хромосомах [32]. Эти данные могут свидетельствать о том, что гены на W-хромосоме могут быть вовлечены в процесс деметилирования МНМ-участка Z-хромосомы. У ZZ-самцов в отсутствие W-хромосомы МНМ-участок остается гипер-метилированным и неактивным, не синтезируется РНК и не происходит подавление активности DMRT1 (рис. 1).
Если предположить, что роль фактора F выполняет продукт гена FET1, то результатом подавления его активности должен быть эффект, наблюдаемый при ингибировании активности ароматазы, т. е. реверсия пола в сторону самца. Нокдаун гена FET1 позволил бы прояснить это предположение, как это было показано с геном DMRT1 [4].
Раздел об определении пола у бабочек и птиц следует начать с небольшого отступления. В самом деле, мы только что выяснили способ определения пола у дрозофилы и у животных вообще и подчеркнули его подкупающую простоту и широкую распространенность в мире животных.
И вот мы снова встречаемся с очередной загадкой природы, с новым усложнением интересующего нас вопроса. Оказывается, все сказанное выше об определении пола по типу дрозофилы правильно, но за одним исключением: этот тип определения пола является в природе не единственным, общим для всех организмов. Наряду с ним существует еще один способ, или тип, определения пола, впервые открытый у бабочек, а затем у птиц и в том числе у домашней курицы. По имени насекомого, на котором этот тип определения пола был открыт впервые, он и носит название типа бабочек. Рассмотрим его особенности и отличия от типа дрозофилы. В качестве объекта для описания процесса возьмем кур: с ними читатель, несомненно, знаком ближе, чем с бабочками; да и в дальнейшем нам еще неоднократно придется иметь с ними дело.
Итак, в чем заключается отличие механизма определения пола у птиц и у дрозофилы?
У дрозофилы, как и у всех животных, самцы продуцируют два типа сперматозоидов - с Х- или У-хромосомой, и в этом смысле они играют решающую роль в определении пола будущих зародышей. У самок же образуется один тип яиц - с Х-хромосомой.
У бабочек и птиц эти отношения диаметрально противоположны: у них привилегия продуцировать два типа половых клеток принадлежит самкам, вследствие чего половина откладываемых ими яиц (на самок) содержит одну половую хромосому, и половина яиц (на самцов) содержит другую, несходную с первой половую хромосому. У самцов же бабочек и птиц образуется один тип сперматозоидов. Следовательно, женский пол у них гетерогаметен, а мужской пол гомогаметен.
Что касается делений созревания яиц и сперматозоидов, то они и здесь протекают так же, как это было описано выше для дрозофилы и человека: первое из них, или собственно редукционное, деление протекает по типу мейоза, и второе, или эквационное, - по типу митоза.
Для того чтобы подчеркнуть разницу в способах определения пола у дрозофилы и у животных, с одной стороны, и у бабочек и птиц, с другой, половые хромосомы последних иногда обозначают другими буквами, а именно - Z и W. Согласно этой системе, половые хромосомы самца обозначают буквами ZZ, а половые хромосомы самки - ZW. Соответственно этому продуцируемый петухом один тип сперматозоидов обозначают буквой Z, а продуцируемые курицей два типа яиц - буквами Z (на самцов) и W (на самок).
Однако, следуя имеющимся в литературе прецедентам, мы отступим от этого правила и в дальнейшем будем придерживаться единой системы обозначения половых хромосом, независимо от того, идет ли речь об определении пола по типу дрозофилы или по типу бабочек и птиц. Дело заключается не в том, какими буквами обозначать половые хромосомы двух сравниваемых групп организмов; важнее помнить, что в отличие от дрозофилы, у которой гетерогаметен мужской пол, у бабочек и птиц гетерогаметен женский пол и что у них пол зародышей устанавливается в процессе созревания яиц, т. е. еще до оплодотворения.
В то же время единая система обозначения половых хромосом для всех представителей мира животных, за исключением отмеченной полярности типов определения пола, несомненно, способствует более целостному и ясному их пониманию.
Поэтому яйца бабочек и птиц на самцов мы будем обозначать в дальнейшем буквой Х, а яйца на самок - буквой У. Что касается сперматозоидов, то здесь они одного типа; их мы будем обозначать буквой Х. Процесс сперматогенеза и овогенеза у бабочек и птиц протекает точно так же, как и у дрозофилы (см. рис. 14).
Дальнейшие детали процесса определения пола у бабочек и птиц столь же просты, как и у дрозифилы, и сводятся к следующему. Если зрелое яйцо, например, курицы содержит Х-хромосому, то из него после оплодотворения Х-сперматозоидом разовьется петушок (ХХ). Если же яйцо содержит У-хромосому, то из него после оплодотворения (таким же сперматозоидом - у петухов все они одинаковы) разовьется курочка (ХУ) (рис. 24).
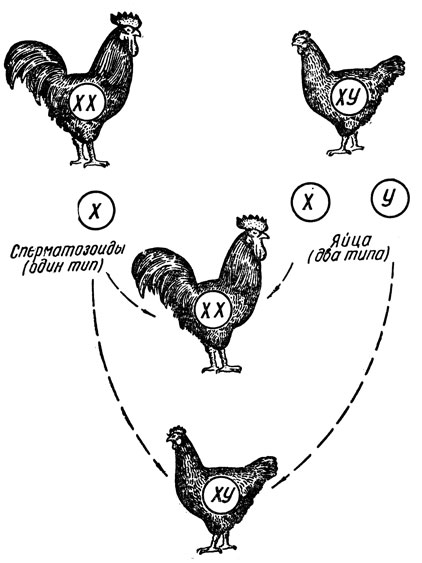
Рис. 24. Определение пола у кур. Вверху изображены петух и курица, в пружках - их половые хромосомы и образующиеся у кур яйца двух типов (на курочек и петушков) и один тип сперматозоидов у петуха. Аутосомы на рисунке не представлены
В соответствии с полярностью механизмов определения пола у дрозофилы и у птиц по-разному представляются и результаты оплодотворения. В самом деле, у дрозофилы, как мы видели, пол зародыша определяется в момент оплодотворения и в каждом отдельном случае зависит от сочетания половых хромосом в оплодотворенном яйце. В отличие от дрозофилы у бабочек и птиц оплодотворение яйца, образно выражаясь, лишь дает толчок развитию зародыша того пола, который уже заложен в нем в процессе созревания. Таким образом, каждому куриному яйцу в буквальном смысле слова "написано на роду" развиться в цыпленка именно того, а не противоположного пола.
Необходимо также иметь в виду, что в клетках птиц и бабочек, как и у всех организмов, кроме половых хромосом, содержатся и наборы аутосом. Диплоидное число хромосом у курицы равно 78. Соответственно этому половина яиц курицы содержит Х-хромосому и 38 аутосом (Х + 38) и половина яиц - У-хромосому и столько же аутосом (У + 38). Сперматозоиды петуха все одинаковы - они содержат Х-хромосому и 38 аутосом (Х + 38).
К сказанному выше об определении пола у кур необходимо сделать следующую оговорку. Дело в том, что ввиду наличия у курицы большого числа очень мелких хромосом и трудностей их подсчета и идентификации вопрос о наличии у нее У-хромосомы до сих пор окончательно не решен, и возможно, что ее здесь нет вообще.
Если в будущем это окажется справедливым, то все сказанное выше об определении пола у кур останется в силе, за тем исключением, что состав половых хромосом курицы нужно будет обозначать как ХО, а продуцируемые ею два типа яиц соответственно как Х + 38 и 0 + 38. Общее же число хромосом будет при этом условии на единицу меньше, т. е. 77. Обозначения половых хромосом петyxa и продуцируемых им сперматозоидов останутся теми же, а диплоидное число хромосом у петуха - на единицу больше, чем у курицы.
Диплоидное число хромосом у бабочек и в том числе у шелкопряда (см. главу IV) равно 56.
22 августа 2018
- 1051
- 0,2
- 1
- 3

Новость
Пятипалый свистун (Leptodactylus pentadactylus)
Автор
Редактор
- Биология
- Вопросы пола
- Генетика
- Цитология
У позвоночных животных очень часто ключевую роль в определении пола играют половые хромосомы. Если у низших позвоночных в определении пола также нередко участвуют факторы окружающей среды, то у птиц и млекопитающих определение пола строго хромосомное. Как правило, в кариотипе есть две половые хромосомы: X и Y у млекопитающих (самки имеют кариотип XX, самцы — XY) или Z и W у птиц (ZW у самок и ZZ у самцов). Впрочем, иногда половых хромосом в кариотипе больше двух. Абсолютным рекордсменом по этому показателю долгое время считался утконос: из 52 его хромосом в качестве половых функционируют 10. Однако недавно невзрачная южноамериканская лягушка, известная как пятипалый свистун (Leptodactylus pentadactylus), уверенно утерла ему нос: из 22 ее хромосом более половины (а именно, 12) являются половыми! Наша статья посвящена этому любопытнейшему открытию.
У многих низших позвоночных — рыб, амфибий и рептилий — как таковых половых хромосом, морфологически отличающихся от остальных хромосом (аутосом), нет. При этом у млекопитающих и птиц обязательно есть половая хромосома , утратившая значительную часть генов — Y-хромосома в случае млекопитающих и W-хромосома в случае птиц. В тех случаях, когда половые хромосомы все-таки есть, они обычно представлены одной парой: XX♀:XY♂ или ZZ♂:ZW♀. Причины, по которым у низших позвоночных нет морфологически выделяющихся (гетероморфных) половых хромосом, не совсем ясны. Существует два предположения на этот счет. Согласно одному из них, мутации в генах, участвующих в определении пола, происходят так часто, что хромосомы просто не имеют возможности начать терять их из-за необходимости все время устранять мутации в этих чрезвычайно важных генах, возвращаясь к исходному состоянию. Вторая гипотеза предполагает, что дегенерации половых хромосом препятствуют многочисленные акты рекомбинации, в ходе которых утерянные фрагменты восстанавливаются.
«Биомолекула» не раз писала о различных аспектах биологии пола. О том, что такое пол с точки зрения биологии, можно прочитать в статьях «Что такое пол?» [1] и «Мальчики налево, девочки направо. А остальные?» [2]. Влиянию условий окружающей среды на формирование пола посвящена статья «Могут ли условия жизни птиц влиять на пол потомства?» [3]. Об особенностях хроматина в ядрах гамет, а также его перестройках при образовании зиготы можно узнать из статьи «Половая жизнь хроматина» [4].
Однако в биологии нет правил без исключений. Известны примеры амфибий, имеющих несколько гетероморфных половых хромосом. Например, у лягушек Strabomantis biporcatus и Pristimantis riveroi определение пола происходит по схеме X1X1X2X2♀:X1X2Y♂. В 2016 году в Швеции была найдена популяция травяных лягушек (Rana temporaria), у которых имеется две Х-хромосомы и две Y-хромосомы. Большинство примеров наличия нескольких половых хромосом относится к млекопитающим. Например, у утконоса имеется 10 половых хромосом, из которых 5 являются Х-хромосомами и 5 — Y-хромосомами [5].

Рисунок 1. Кольцевая структура, формирующаяся при мейозе у самцов пятипалого свистуна. Отчетливо видны 12 хромосом, образующих кольцо. ДНК окрашена синим, красным выделены теломеры.
[5], рисунок с изменениями
Как известно, при кроссинговере гомологичные хромосомы сцепляются друг с другом с образованием бивалентов. У человека Y-хромосома потеряла так много генов, что участков гомологии с почти не осталось, поэтому рекомбинации между этими хромосомами практически не происходит. Впрочем, у многих видов различия между половыми хромосомами не столь значительны, и при кроссинговере они, как и аутосомы, соединяются с образованием бивалента. Однако как ведут себя при мейозе несколько половых хромосом? У самцов, имеющих несколько Х-хромосом и несколько Y-хромосом, образуются не биваленты, а особые кольцевые структуры, в которых каждая хромосома взаимодействует с гомологичными участками двух своих соседей. Это возможно благодаря тому, что на всех X- и Y-хромосомах имеются гомологичные псевдоаутосомные районы. На финальных этапах мейоза кольцевая структура упорядоченно разделяется, и сперматозоиды получают либо n Х-хромосом, либо n Y-хромосом, где n — число хромосом в гаплоидном хромосомном наборе данного вида. Следовательно, наличие характерной кольцевой структуры в мейозе клеток-предшественников сперматозоидов является верным признаком наличия у вида нескольких половых хромосом.
Именно такую структуру удалось обнаружить в сперматоцитах пятипалого свистуна с помощью флуоресцентной микроскопии (рис. 1). Она видна и на препаратах хромосом, окрашенных по Гимзе (рис. 2) [5].
О том, что такое флуоресцентная микроскопия и как она работает, можно прочитать в статье «12 методов в картинках: микроскопия» [6].

Рисунок 2. Кольцевые структуры в сперматоцитах двух самцов пятипалого свистуна. Хромосомы окрашены по Гимзе. Масштабная линейка 5 мкм.
[5], рисунок с изменениями
Кольцевая структура в сперматоцитах пятипалого свистуна состоит из 12 хромосом, при этом полный кариотип этой лягушки включает 22 хромосомы. Таким образом, пятипалый свистун — единственный известный на данный момент вид позвоночных, в кариотипе которого половых хромосом больше, чем аутосом. Ученые предполагают, что Y-хромосома пятипалого свистуна претерпела целых семь транслокаций, которые и привели к такому большому количеству половых хромосом. Однако причины, лежащие в основе таких значительных перестроек, неясны.
Случай пятипалого свистуна, конечно, является уникальным для амфибий и позвоночных вообще. Однако он прекрасно иллюстрирует всю сложность и многообразие генетических основ определения пола.
Читайте также: